Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Antecedentes históricos
Co-linearidad entre el ADN y la proteína codificada por ese ADN. Yanofsky, demostró que el orden de ciertas mutaciones en el gen de la triptofano sintetasa era el mismo al de los cambios de aminoácidos en la proteína Crick y Brenner a partir de una larga colección de doble mutantes de T4, que el código genético es leído en forma secuencial a partir de un punto fijo

2
Estos experimentos sólo indican una correlación
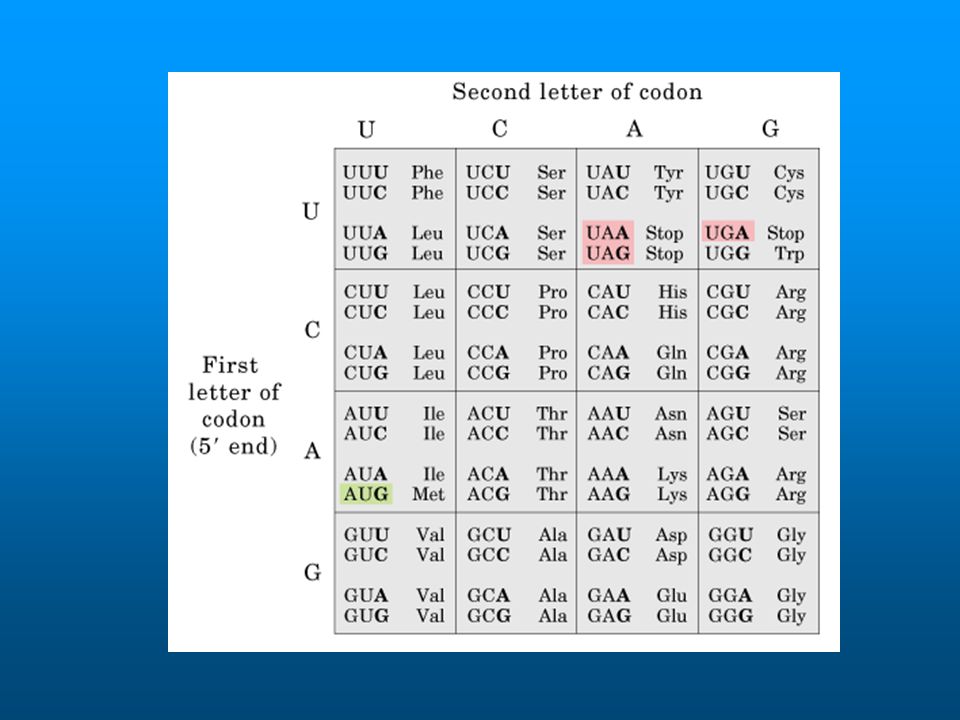
El diccionario preciso del código genético se determinó utilizando sistemas de traducción in vitro derivado de células de E.coli

6
Har Gobind Khorana

10
PREFERRED CODONS FOR SELECTED SPECIES
The following table lists the preferred codons in each species, along with their usage as a percent of all codons for that amino acid. Codons that differ from those preferred in man and rat are highlighted Amino acid Human Rat E. coli S. cerevisiae S. frugiperda Preferred codon % use Ala GCC 41 34 GCT 38 37 Arg CGG 21 AGG CGC AGA 48 24 Asn AAC 55 60 54 AAT 59 63 Asp GAC 58 GAT 65 Cys TGC 56 TGT Gln CAG 75 76 66 CAA 69 51 Glu GAG 62 GAA 68 71 52 Gly GGC 35 39 GGT 47 GGA 32 His CAC CAT 57 64 Ile ATC 50 ATT 46 Leu CTG 42 49 TTG 29 31 Lys AAG AAA Met ATG 100 Phe TTC TTT Pro CCC 33 CCG CCA CCT/CCA Ser AGC 25 26 TCT 27 TCC 20 Thr ACC ACT Trp TGG Tyr TAC 61 TAT 67 Val GTG 36 GTT Trm TGA TAA

11
“La molécula adaptadora”

12
A partir de la secuencia de 300 tRNAs

13
La Hipótesis del Balanceo (The Wobble Hypothesis)
Las células contienen diferentes tRNAs que son específicos para el mismo aminoácido Muchos tRNAs unen a 2 o 3 codones

14
Activación de los aminoácidos
Al menos 20 aminoacil-tRNA sintetasas La activación requiere ATP La especificidad no es determinada por el anticodón Proof read antes de liberar el producto

15
X174

18
Iniciación Se requiere un tRNA específico (tRNAmeti)
En E.coli una vez unida la metionina se formila En ecuariotas tRNAmeti es específico pero no es formilado El codón de iniciación (AUG) En procariotas es localizado adyacente al elemento Shine-Dalgarno En ecuariotas es GENERALMENTE el primer AUG encontrado por el ribosoma (A/G CCA/G CC AUG A/G)
 En procariotas es localizado adyacente al elemento Shine-Dalgarno. En ecuariotas es GENERALMENTE el primer AUG encontrado por el ribosoma (A/G CCA/G CC AUG A/G)")
19
Secuencia de reconocimiento de la iniciación de la traducción

20
Small GTP-binding proteins require helper proteins, to
facilitate GDP/GTP exchange, or promote GTP hydrolysis. A guanine nucleotide exchange factor (GEF) induces a conformational change that makes the nucleotide-binding site of a GTP-binding protein more accessible to the aqueous intracellular milieu, where [GTP] [GDP]. Thus a GEF causes a GTP-binding protein to release GDP & bind GTP (GDP/GTP exchange).
 induces a conformational change that makes the nucleotide-binding site of a GTP-binding protein more accessible to the aqueous intracellular milieu, where [GTP] [GDP]. Thus a GEF causes a GTP-binding protein to release GDP & bind GTP (GDP/GTP exchange).")
21
A GTPase activating protein (GAP) causes a GTP-binding protein to hydrolyze its bound GTP to GDP + Pi. The active site for GTP hydrolysis is on the GTP-binding protein, although a GAP may contribute an essential active site residue. GEFs & GAPs may be separately regulated. Unique GEFs and GAPs interact with different GTP-binding proteins

22
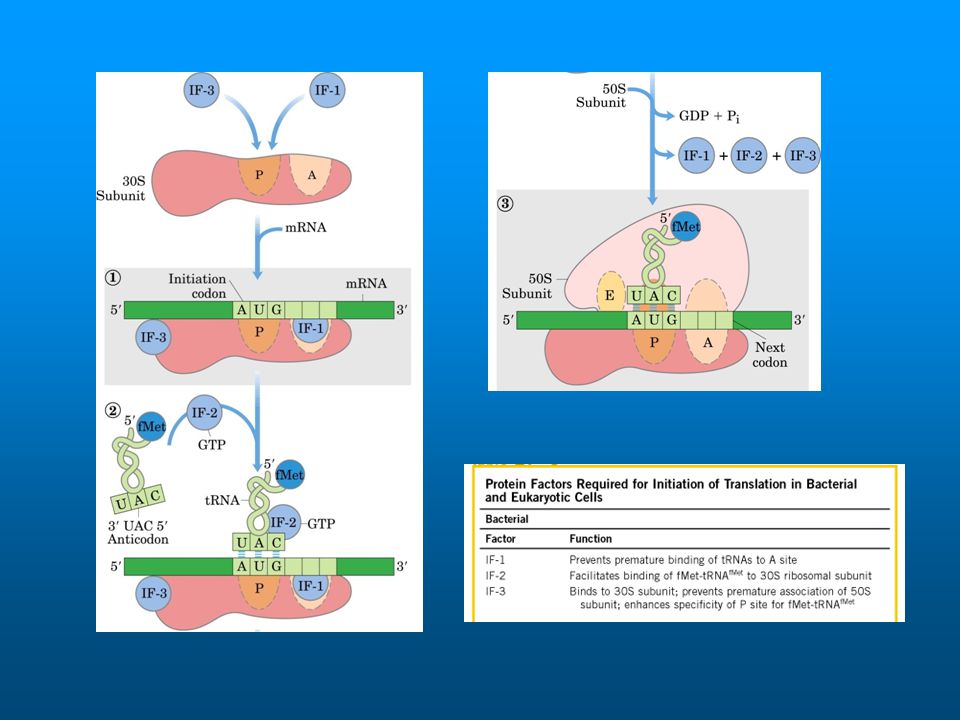
Initiation of protein synthesis in E
Initiation of protein synthesis in E. coli requires initiation factors IF-1, IF-2, & IF-3. IF-3 binds to the 30S ribosomal subunit, freeing it from its complex with the 50S subunit. IF-1 assists binding of IF-3 to the 30S ribosomal subunit. IF-1 also occludes the A site of the small ribosomal subunit, helping insure that the initiation aa-tRNA fMet-tRNAfMet can bind only in the P site & that no other aa-tRNA can bind in the A site during initiation. IF-2 is a small GTP-binding protein. IF-2-GTP binds the initiator fMet-tRNAfMet & helps it to dock with the small ribosome subunit.

24
As the large ribosomal subunit joins the complex,
As mRNA binds, IF-3 helps to correctly position the complex such that the tRNAfMet interacts via base pairing with the mRNA initiation codon (AUG). A region of mRNA upstream of the initiation codon, the Shine-Dalgarno sequence, base pairs with the 3' end of the 16S rRNA. This positions the 30S ribosomal subunit in relation to the initiation codon. As the large ribosomal subunit joins the complex, GTP on IF-2 is hydrolyzed, leading to dissociation of IF-2-GDP and dissociation of IF-1. A domain of the large ribosomal subunit serves as GAP (GTPase activating protein) for IF-2. Once the two ribosomal subunits come together, the mRNA is threaded through a curved channel that wraps around the "neck" region of the small subunit.
. A region of mRNA upstream of the initiation codon, the Shine-Dalgarno sequence, base pairs with the 3 end of the 16S rRNA. This positions the 30S ribosomal subunit in relation to the initiation codon. As the large ribosomal subunit joins the complex, GTP on IF-2 is hydrolyzed, leading to dissociation of IF-2-GDP and dissociation of IF-1. A domain of the large ribosomal subunit serves as GAP (GTPase activating protein) for IF-2. Once the two ribosomal subunits come together, the mRNA is threaded through a curved channel that wraps around the neck region of the small subunit.")
26
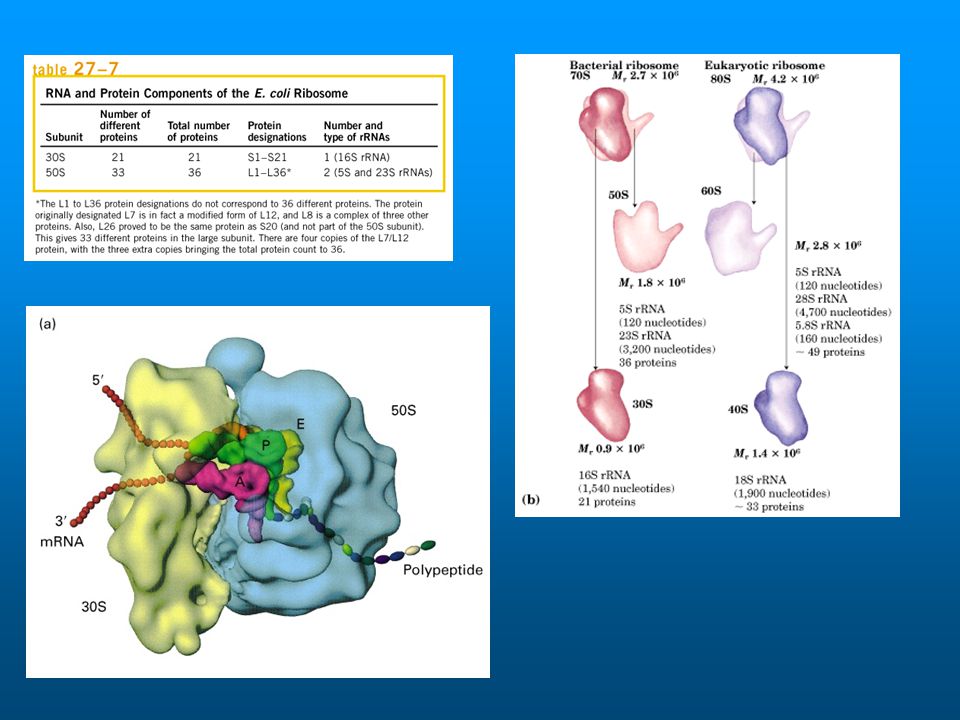
Elongation cycle Ribosome structure and position of factors & tRNAs based on cryo-EM with 3D image reconstruction. Diagram provided by Dr. J. Frank, Wadsworth Center, NYS Dept. of Health. Partial images on subsequent slides are derived from this. Colors: large ribosome subunit, cyan; small subunit, pale yellow; EF-Tu, red; EF-G, blue. tRNAs, gray, magenta, green, yellow, brown.

27
Elongation requires participation of elongation factors
EF-Tu (also called EF1A) EF-Ts (EF1B) EF-G (EF2) EF-Tu & EF-G are small GTP-binding proteins. The sequence of events follows.
 EF-Ts (EF1B) EF-G (EF2) EF-Tu & EF-G are small GTP-binding proteins. The sequence of events follows.")
28
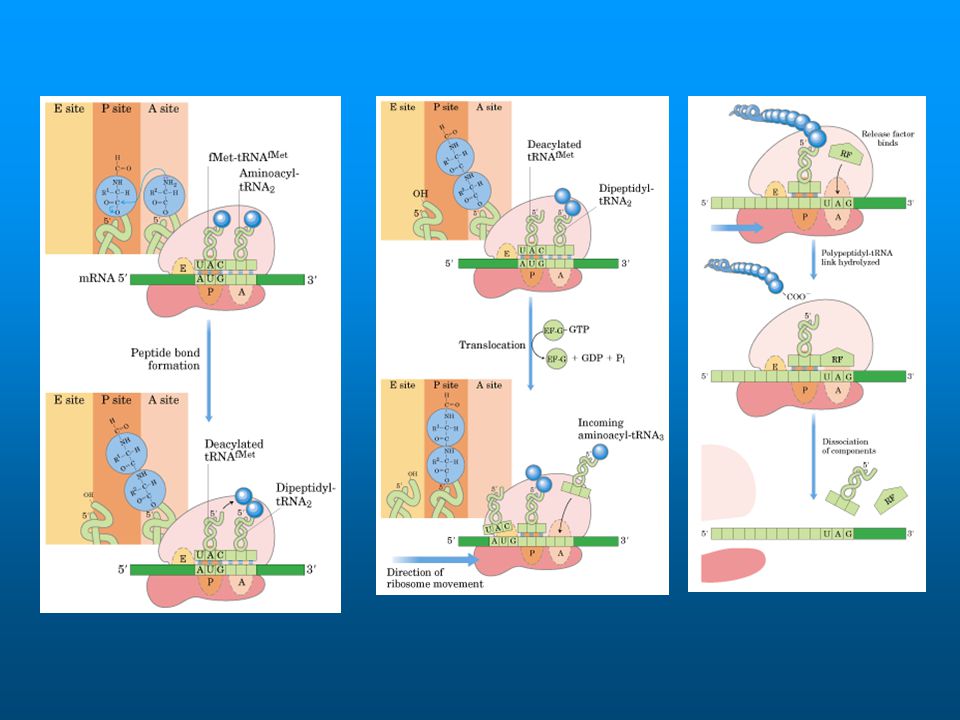
EF-Tu-GTP binds & delivers an aminoacyl-tRNA to the A site
on the ribosome. EF-Tu recognizes & binds all aminoacyl-tRNAs with approx. the same affinity, when each tRNA is bonded to the correct (cognate) amino acid. tRNAs for different amino acids have evolved to differ slightly in structure, to compensate for different binding affinities of amino acid side-chains, so the aminoacyl-tRNAs all have similar affinity for EF-Tu. EF-Tu colored red
 amino acid. tRNAs for different amino acids. have evolved to differ slightly. in structure, to compensate for. different binding affinities of amino acid side-chains, so the aminoacyl-tRNAs all have similar affinity for EF-Tu. EF-Tu colored red.")
29
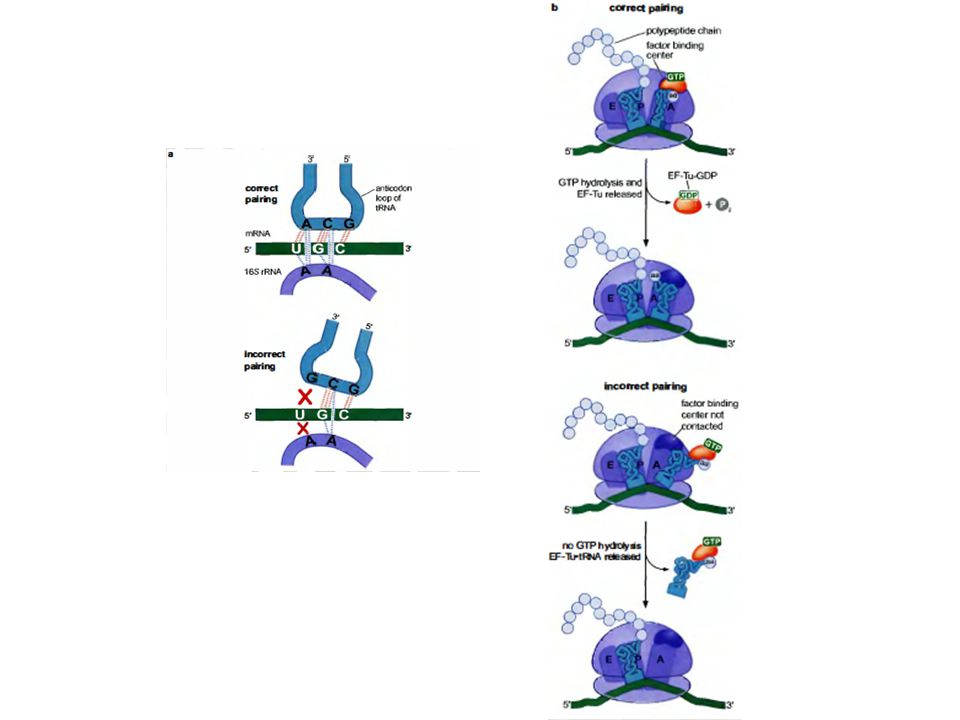
The tRNA must have the correct anticodon to interact with the mRNA codon positioned at the A site to form a base pair of appropriate geometry. Universally conserved bases of 16S rRNA interact with and sense the configuration of the minor groove of the short stretch of double helix formed from the first 2 base pairs of the codon/anticodon complex. A particular ribosomal conformation is stabilized by this interaction, providing a mechanism for detecting whether the correct tRNA has bound. Proofreading in part involves release of the aminoacyl-tRNA prior to peptide bond formation, if the appropriate ribosomal conformation is not generated by this interaction.

31
The change in ribosomal conformational associated with formation of a correct codon-anticodon complex leads to altered positions of active site residues in the bound EF-Tu, with activation of EF-Tu GTPase activity. The ribosome thus functions as GAP for EF-Tu.

32
ribosome, the tRNA initially has a distorted conformation.
When EF-Tu delivers an aminoacyl-tRNA to the ribosome, the tRNA initially has a distorted conformation. As GTP on EF-Tu is hydrolyzed to GDP + Pi , EF-Tu undergoes a large conformational change & dissociates from the complex. The tRNA conformation relaxes, & the acceptor stem is repositioned to promote peptide bond formation. This process is called accommodation. EF-Tu colored red

33
It includes rotation of the single-stranded 3' end of the acceptor stem of the A-site tRNA around an axis that bisects the peptidyl transferase center of the ribosomal large subunit. This positions the 3' end with its attached amino acid in the active site, near the 3' end of the P-site tRNA, & adjacent to the mouth of the tunnel through which nascent poly-peptides exit the ribosome.

34
EF-Ts functions as GEF to reactivate EF-Tu.
Interaction with EF-Ts causes EF-Tu to release GDP. Upon dissociation of EF-Ts, EF-Tu binds GTP, which is present in the cytosol at higher concentration than GDP.

35
Transpeptidation (peptide bond formation) involves nucleophilic attack of the amino N of the amino acid linked to the 3'OH of the terminal adenosine of the tRNA in the A site on the carbonyl C of the amino acid (with attached nascent polypeptide) in ester linkage to the tRNA in the P site. The reaction is promoted by the geometry of the active site consisting solely of residues of the 23S rRNA of the large ribosomal subunit. No protein is found at the active site.

36
The 23S rRNA may be considered a "ribozyme."
As part of the reaction a proton (H+) is extracted from the attacking amino N.
 is extracted from the attacking amino N.")
37
This H+ is then donated to the hydroxyl of the tRNA in the P site, as the ester linkage is cleaved.

38
The nascent polypeptide, one residue longer, is now linked to the A-site tRNA.

39
tRNA grey, EF-Tu red, EF-G blue
The unloaded tRNA in the P site will shift to the E (exit) site during translocation. Translocation of the ribosome relative to mRNA involves the GTP-binding protein EF-G. The size & shape of EF-G are comparable to that of the complex of EF-Tu with an aa-tRNA. Structural studies & molecular dynamics indicate that EF-G-GTP binding in the vicinity of the A site causes a ratchet-like motion of the small ribosomal subunit against the large subunit.
 site during translocation. Translocation of the ribosome relative to mRNA involves the GTP-binding protein EF-G. The size & shape of EF-G are comparable to that of the complex of EF-Tu with an aa-tRNA. Structural studies & molecular dynamics indicate that EF-G-GTP binding in the vicinity of the A site causes a ratchet-like motion of the small ribosomal subunit against the large subunit.")
40
The tRNA with attached nascent polypeptide is pushed from the A site to the P site.
Unloaded tRNA that was in the P site shifts to the E site. Since tRNAs are linked to mRNA by codon-anticodon base pairing, the mRNA moves relative to the ribosome.

41
En E.coli hay 2 RFs, en eucariotas 1 RF
UAA y UAG son reconocidos por RF-1 UAA y UGA son reconocidos por RF-2

42
En eucariotes Iniciación

44
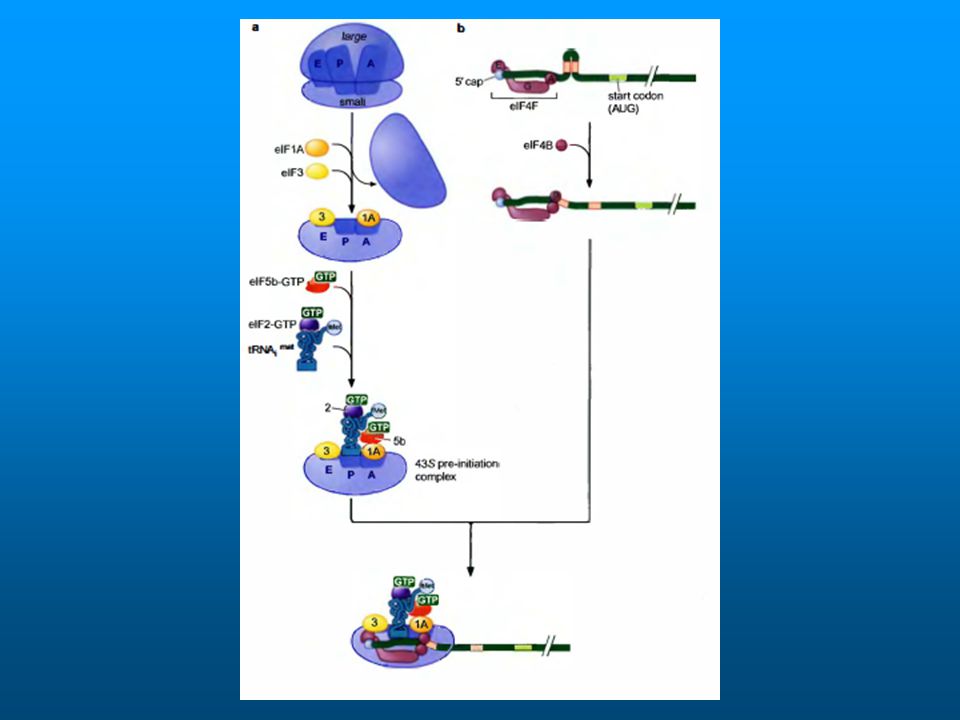
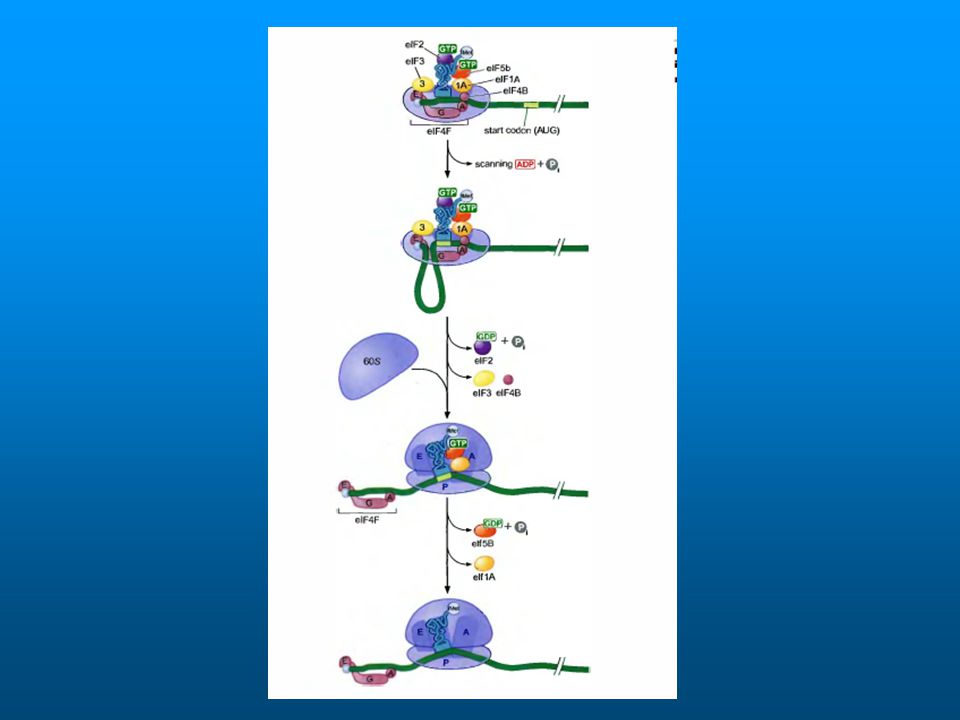
El primer paso es la formación del complejo GTP – eIF-2
eIF-GTP se une a tRNA i Se une a la subunidad 40S (complejo 43S) Luego se estabiliza con eIF-3 y eIF-1 El cap se une a eIF-4F: compuesto por 4E, A y G 4E se une a cap 4A une ATP y tiene actividad RNA helicasa 4G ayuda a la unión al complejo 43 S
 Luego se estabiliza con eIF-3 y eIF-1. El cap se une a eIF-4F: compuesto por 4E, A y G. 4E se une a cap. 4A une ATP y tiene actividad RNA helicasa. 4G ayuda a la unión al complejo 43 S.")
48
Regulación de la actividad eIF-4E
Hipótesis Regulación de la actividad eIF-4E eIF-4E es el IF en menor nivel (determinante) Se regula a nivel de Transcripción Modificación (fosforilación ) Inhibición
 Se regula a nivel de. Transcripción. Modificación (fosforilación ) Inhibición.")
49
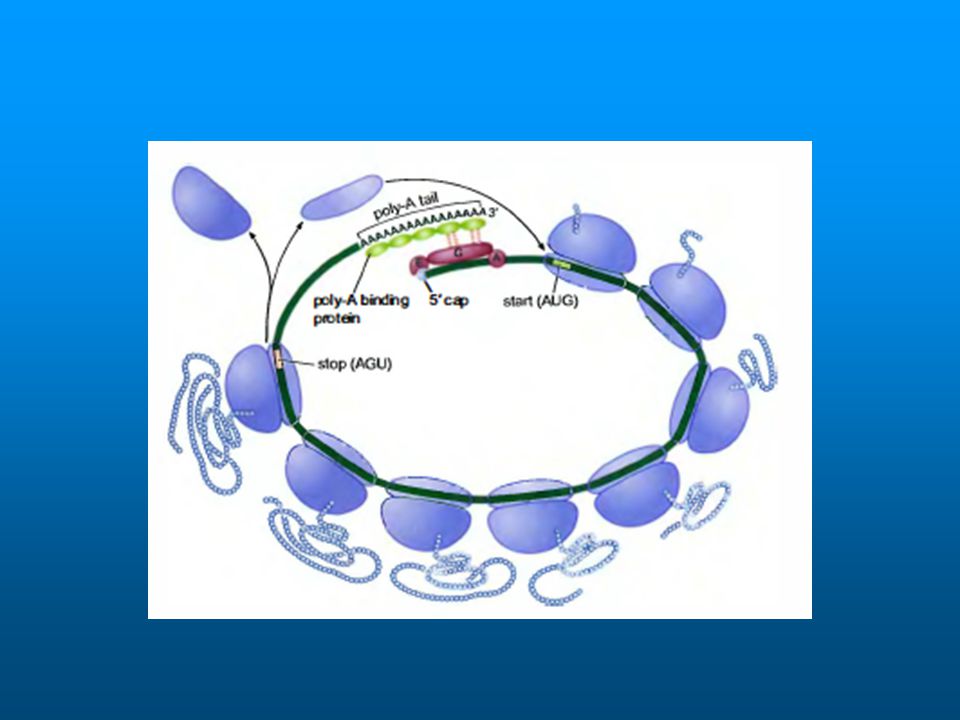
Regulation of Step 1 1. Phosphorylation of the eIF4E Binding Proteins, the 4E-BPs. 2. Binding of PolyAdenylate Binding Protein (PABP) to eIF4G. Why? Because this circularizes the polysome, and allows ribosomal subunits to start new ribosomes.

50
MAPK-Dependent Phosphorylation of eIF4E
Is Mediated by the eIF4G Associated Kinase Mnk vía MAPK/ERK o vía MAPK (de las siglas en inglés Mitogen-activated protein kinases Phosphorylation of eIF4E allows it to detach from the cap and recycle

51
Binding of PolyAdenylate Binding Protein (PABP) to eIF4G
The translational control of maternally inherited mRNAs is a key feature of early animal development mRNAs are synthesized and stored (i.e., masked) during the protracted period of oogenesis and are translated only in response to subsequent exogenous cues the oocytes of Xenopus laevis contain masked mRNAs that are activated during the cell’s reentry into meiosis(oocyte maturation), a process that is stimulated by the interaction of progesterone with a surface-associated receptor
 during the protracted period of oogenesis and are translated only in response to subsequent exogenous cues. the oocytes of Xenopus laevis contain masked mRNAs that are activated during the cell’s reentry into meiosis(oocyte maturation), a process that is stimulated by the interaction of progesterone with a surface-associated receptor.")
53
Polyadenylation Leads to the PABP-mediated Displacement of Maskin from eIF4E
In case of mRNAs with a CPE sequence in the 3’ end, the poly(A) tail also serves to disrupt the binding of maskin, a CPEB-binding protein to eIF4E. This makes eIF4E available to start building the cap-binding complex
 tail also serves to disrupt the binding of maskin, a CPEB-binding protein to eIF4E. This makes eIF4E available to start building the cap-binding complex.")
54
Control de la síntesis de Hemo por traducción
Regulación de la iniciación

55
Control de la síntesis de Hemo por traducción
Protein synthesis in intact reticulocytes and their lysates is dependent on the availability of heme. In heme deficiency, protein synthesis is inhibited with increased phosphorylation of the a subunit of eIF2. The phosphorylation of eIF2 in heme deficiency is the result of the activation of heme-regulated inhibitor (HRI), which is a heme-regulated eIF2 kinase
, which is a heme-regulated eIF2 kinase.")
56
eIF-2 GTP GDP GEF (eIF-2B) P HRI inactivo activo Alto hemo Bajo hemo
 P HRI inactivo activo Alto hemo Bajo hemo")
57
Elongación Amino acid-containing tRNA molecules (aminoacyl-tRNAs, aa-tRNA) are picked up by elongation factor eEF-1 in the presence of GTP. The empty tRNA is displaced from the P-site to the E-site as the peptidyl tRNA is translocated from the A-site to the P-site. The process is facilitated by elongation factor eEF-2 and GTP.

58
Terminación Releasing Factors (eRFs) are involved in termination.
eRF1 structurally mimics tRNA that is bound to eEF1a • GTP. eRF1 fits into the ribosomal A-site, where it recognizes the stop codon. It then releases the completed polypeptide by catalyzing a nucleophilic attack on the ester bond between the peptide and the P-site tRNA. The catalytic activity of eRF1 is stimulated by the GTP-bound form of another relasing factor, eRF3.

59
Mimicry of tRNA by the Releasing Factor eRF1
Domain 2 on eRF1 Recognizes the Stop Codon Domain 3 Catalyzes the Hydrolysis of the Completed Peptide from the P-Site tRNA

60
Se une a la subunidad 30S del ribosoma bacteriano bloqueando la fijación del aminoacil-tRNA al sitio aceptor (A) del complejo formado por el mRNA y la subunidad 50S del ribosoma
 del complejo formado por el mRNA y la subunidad 50S del ribosoma")
61
La kanamicina afecta a la subunidad 30S de los ribosomas y causa una mutación con cambio previniendo la traducción del ARN. Aminoacil- tARN, en vez de leer el codón CUA —como sería el caso en la secuencia CUAG— se lee el codón UAG (codón ambar de terminación)
")
62
Translational Frameshift

63
terminación de la traducción Frameshift
UGA C UGA GAC RF2 RF2 Pausa Unión de RF2 terminación de la traducción Frameshift (UGAC se lee como GAC y sigue +1)
")
64
Experimento de Dintzis
Determinación de la dirección de síntesis de las proteínas Experimento de Dintzis

65
Ingram’s fingerprinting technique was performed by purifying hemoglobin from red blood cells, fragmenting hemoglobin protein into peptides with the enzyme trypsin, separating the fragments (based on their respective charges) by electrophoresis, and staining his results. In this way he produced a “fingerprint” of the protein, as the different amino acids would migrate to different locations on the electrophoretic gradient based on their charges. Dintzis added 14C-labeled amino acids to mature reticulocytes, which are always involved in synthesizing hemoglobin.

67
¿Qué resultados se obtienen?
¿Qué se grafica?

Presentaciones similares











 Hacer: to make/do.>")






>")









