Descargar la presentación
La descarga está en progreso. Por favor, espere

1
¿ Cómo se origina la diversidad en las moléculas de reconocimiento ?
Eficacia del S. Inmune Capacidad de reconocer antígenos Entender el proceso que se discutió en la lámina anterior implicó descifrar importantes enigmas. Por ejemplo como el sistema inmune adaptativo podía generar y seleccionar anticuerpos que reaccionaban contra antígenos que no habían sido enfrentados en el curso de la evolución de estos organismos (por ejemplo cuando los antígenos eran nuevos compuestos producidos por síntesis química). Inicialmente se postuló que existía un repertorio variado de receptores y que estos se seleccionaban de algún modo para ser producidos y reaccionar con el antígeno (teoría selectiva), mientras que otros investigadores proponían la hipótesis de que los anticuerpos tenían la capacidad de adaptar su reconocimiento al antígeno en cuestión (teoría instructiva). Ambas implicaban importantes desafíos, en el primer caso debido a la dificultad para concebir como la enorme diversidad requerida podía estar disponible en la información genética del organismo, mientras que en el segundo la dificultad tenía que ver con como una misma proteína podía “cambiar” su estructura para reaccionar con antígenos muy diferentes. Al comenzar a secuenciarse anticuerpos de mielomas (tumor maligno derivado de células de la medula osea, que hace que algunos pacientes tengan en el plasma enormes cantidades de un único anticuerpo, o en oportunidades la cadena liviana del mismo, secretados por el tumor) se vió que los distintos mielomas tenían distintas secuencias, descartando la teoría instructiva Respecto a la dificultad para comprender como era posible la enorme información genética que requería la teoría selectiva si se asumía la idea imperante en los sesenta de “un gen una proteína”, en 1965 se propuso que el volumen de información podía reducirse si existiera un mecanismo “modular” por el cual los genes que codificaban la región variable de los anticuerpos, pudiera combinarse con una región constatne común.
. Inicialmente se postuló que existía un repertorio variado de receptores y que estos se seleccionaban de algún modo para ser producidos y reaccionar con el antígeno (teoría selectiva), mientras que otros investigadores proponían la hipótesis de que los anticuerpos tenían la capacidad de adaptar su reconocimiento al antígeno en cuestión (teoría instructiva). Ambas implicaban importantes desafíos, en el primer caso debido a la dificultad para concebir como la enorme diversidad requerida podía estar disponible en la información genética del organismo, mientras que en el segundo la dificultad tenía que ver con como una misma proteína podía cambiar su estructura para reaccionar con antígenos muy diferentes. Al comenzar a secuenciarse anticuerpos de mielomas (tumor maligno derivado de células de la medula osea, que hace que algunos pacientes tengan en el plasma enormes cantidades de un único anticuerpo, o en oportunidades la cadena liviana del mismo, secretados por el tumor) se vió que los distintos mielomas tenían distintas secuencias, descartando la teoría instructiva. Respecto a la dificultad para comprender como era posible la enorme información genética que requería la teoría selectiva si se asumía la idea imperante en los sesenta de un gen una proteína , en 1965 se propuso que el volumen de información podía reducirse si existiera un mecanismo modular por el cual los genes que codificaban la región variable de los anticuerpos, pudiera combinarse con una región constatne común.")
2
¿CÓMO ES POSIBLE QUE SE GENERE SEMEJANTE DIVERSIDAD?

3
Proliferación y diferenciación inducida por el antígeno
Funciona sobre la base de la selección clonal lo cual requiere la generación de una alta diversidad de receptores Proliferación y diferenciación inducida por el antígeno Células de memoria Células efectoras Progenitor común Generación de linfocitos vírgenes con especificidad aleatoria Diversidad:

4
Maduración linfocítica
Comprende: Expresión secuencial de genes produciéndose la proliferación celular. Generación repertorio diverso.(expresión de receptores, f reconocimiento). Cambios fenotípicos de las células (caracterización). Adquisición competencia funcional (f. efectora). Selección linfocitos útiles.
. Cambios fenotípicos de las células (caracterización). Adquisición competencia funcional (f. efectora). Selección linfocitos útiles.")
5
Estadios de la maduración
Maduración precoz: elevada actividad mitótica por acción IL7 de c. estroma de M.Osea y Timo -diferenciación y proliferación. Expresión genes receptores: Dan señales que promueven la supervivencia, proliferación y generación repertorio linfocítico maduro. mecanismo control de calidad. Selección repertorio maduro: S. positiva: conservación de receptores con especificidades útiles que reconocen ags extraños y sin avidez cph. S. negativa: Eliminación de linfocitos de alta avidez cph propio, mecanismo de mantención de la tolerancia.

6
Adquisición de competencia funcional
Fase tardía en que se adquiere: Capacidad de respuesta a antígenos, con generación de mecanismos efectores. Capacidad de secreción de inmunoglobulinas. Desarrollo de diferentes tipos de linfocitos.

7
El proceso de recombinación somática es central en el funcionamiento del sistema inmune adaptativo
Porque es el mecanismo básico de generación diversidad, que produce el repertorio inicial de anticuerpos. 2. Porque el fenómeno de exclusión alélica, genera linfocitos monespecíficos, lo cual es necesario para que ocurra el proceso de selección clonal. Además, de ser el proceso de generación de diversidad y de producción de linfocitos monoespecíficos, durante el proceso de recombinación es un importante evento para la regulación de la transcripción de los genes recombinados. El rearreglo de genes aproxima el promotor que se encuentra en el intrón 5’ del segmento Vn con el potenciador (enhancer) que está en el intrón 5’ de los exones de la región constante promoviendo la transcripción. La acción de los factores de transcripción sobre el enhancer aumenta enormemente la velocidad de transcripción, pero para que ocurra, debe estar físicamente próximo al promotor correspondiente, lo cual ocurre al producirse la recombianción.
 que está en el intrón 5’ de los exones de la región constante promoviendo la transcripción. La acción de los factores de transcripción sobre el enhancer aumenta enormemente la velocidad de transcripción, pero para que ocurra, debe estar físicamente próximo al promotor correspondiente, lo cual ocurre al producirse la recombianción.")
8
Generación genes de receptores
Por “Recombinación somática” de secuencias de DNA heredados, que se encuentran separados y se unen mediante deleción del dna intermediario y empalme de fragmentos restantes, la unión de diferentes segmentos determina el repertorio y se lleva a cabo independiente de la presencia de antígenos.

9
Generación de la diversidad
Diferentes genes para distintos segmentos de las inmunoglobulinas. Cada segmento posee múltiples versiones para regiones variables. Los genes se reordenan al azar en el momento de la síntesis ”diversidad de combinación”. “Diversidad de unión”. Adición y eliminación de nucleótidos por enzima, cambio patrón de lectura. La existencia de la Hipermutación en el caso de linfocitos B.

10
BCR e Igs

11
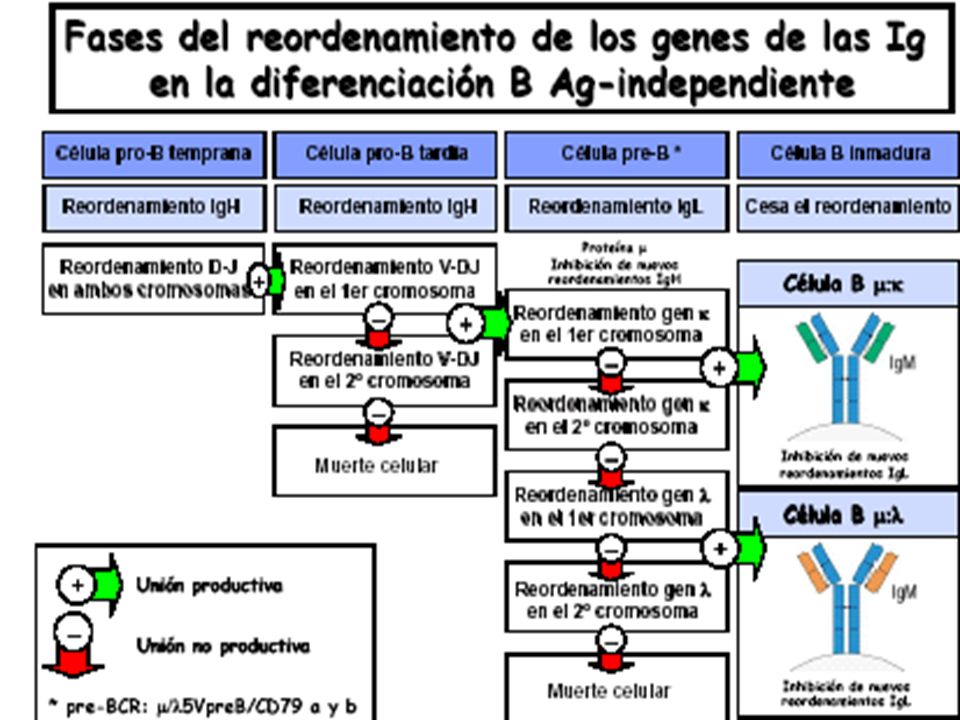
Desarrollo de linfocitos B
No hay expresión de Ig Pro-linfocito B Rearreglo del gen de la cadena pesada Expresión de cadena m + sustituto de cadena liviana Pre-linfocito B Medula ósea Rearreglo del gen de la cadena liviana Linfocito B maduro Co-expresión de IgM e IgD Estímulo por el antígeno El proceso de recombinación somática que hemos visto hasta acá ocurre en la médula osea, y resulta en la expresión del BCR (inmunoglobulina de membrana, junto con las moléculas accesorias α y β). La inmunoglobulina de superficie en este momento es una IgM y si en estas condiciones la especificidad aleatoria que se generó en el linfocito hace que esta interaccione con antígenos propios presentes en la médula, entonces el linfocito B puede recibir señales para reactivar la actividad de las recombinasas RAG 1 y 2, y “reeditar” la cadena liviana. En este proceso, nuevos segmentos génicos V (hacia el 5’ del segmento usado inicialmente) y J (hacia el 3’ del segmento utilizado originalmente) pueden ser usados realizándose un nuevo proceso de recombinación para producir una nueva cadena liviana que al combinarse con la cadena pesada original de la célula en desarrollo puede revertir el reconocimiento del antígeno propio. En caso de no haber reconocimiento de antígenos propios, el linfocito B sigue su desarrollo pasando a ser un linfocito B maduro que por splicing alternativo del RNA primario, coexpresa IgM e IgD en su superficie. Este linfocito está ahora en condiciones de ser exportado a la periferia donde en los correspondientes órganos linfoides puede encontrarse con su antígeno específico. El eventual encuentro con el antígeno y en muchos casos la colaboración con las células T que se verá más adelante en el curso, hará que el linfocito B se active, prolifere y se diferencie. En este proceso ocurrirán nuevos cambios que afectarán la afinidad con que el receptor de superficie del linfocito reconoce el antígeno (maduración de afinidad) y cambios en los exones que codifican para la cadena pesada (cambio de clase) que afectarán la funcionalidad de los anticuerpos secretados Linfocito B activado Tejido linfoide periférico Plasmocito Linfocito B de memoria
. La inmunoglobulina de superficie en este momento es una IgM y si en estas condiciones la especificidad aleatoria que se generó en el linfocito hace que esta interaccione con antígenos propios presentes en la médula, entonces el linfocito B puede recibir señales para reactivar la actividad de las recombinasas RAG 1 y 2, y reeditar la cadena liviana. En este proceso, nuevos segmentos génicos V (hacia el 5’ del segmento usado inicialmente) y J (hacia el 3’ del segmento utilizado originalmente) pueden ser usados realizándose un nuevo proceso de recombinación para producir una nueva cadena liviana que al combinarse con la cadena pesada original de la célula en desarrollo puede revertir el reconocimiento del antígeno propio. En caso de no haber reconocimiento de antígenos propios, el linfocito B sigue su desarrollo pasando a ser un linfocito B maduro que por splicing alternativo del RNA primario, coexpresa IgM e IgD en su superficie. Este linfocito está ahora en condiciones de ser exportado a la periferia donde en los correspondientes órganos linfoides puede encontrarse con su antígeno específico. El eventual encuentro con el antígeno y en muchos casos la colaboración con las células T que se verá más adelante en el curso, hará que el linfocito B se active, prolifere y se diferencie. En este proceso ocurrirán nuevos cambios que afectarán la afinidad con que el receptor de superficie del linfocito reconoce el antígeno (maduración de afinidad) y cambios en los exones que codifican para la cadena pesada (cambio de clase) que afectarán la funcionalidad de los anticuerpos secretados. Linfocito B activado. Tejido linfoide periférico. Plasmocito. Linfocito B de memoria.")
13
Mecanismos de generación de diversidad del BCR
Recombinación somática Diversidad de unión Asociación segmentos al azar Hipermutación somática

14
Síntesis de las cadenas de Igs
Genes que codifican los aa para la cadena pesada. Región constante por gen C, con 9 versiones, cada una para cada isotipo de inmunoglobulina. Región variable codificada por gen V con 3 segmentos génicos V, D, J.

15
Mecanismos de generación de diversidad del BCR
Elección de una versión de cada grupo de genes que codifican las distintas zonas de las cadenas pesadas y livianas de las Igs. Características: Selección al azar de los segmentos. Se produce en forma secuencial. 1º DJ y después V de la cadena pesada 2º VJ y después C de la cadena ligera

16
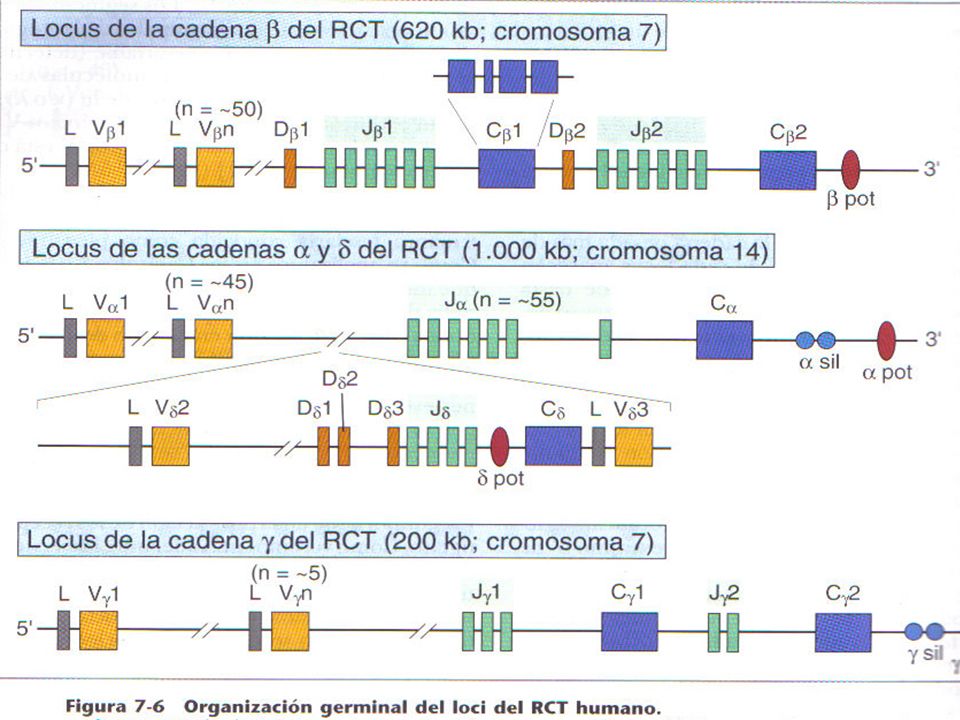
Cadenas Pesadas de Igs Gen para cadenas pesadas H con sus región variable VH y constante CH esta con 9 segmentos génicos. Región V posee segmentos de genes de la región , que son: V, D, J que codifican los aa

17
Organización multigénica de los genes de las Inmunoglobulinas

18
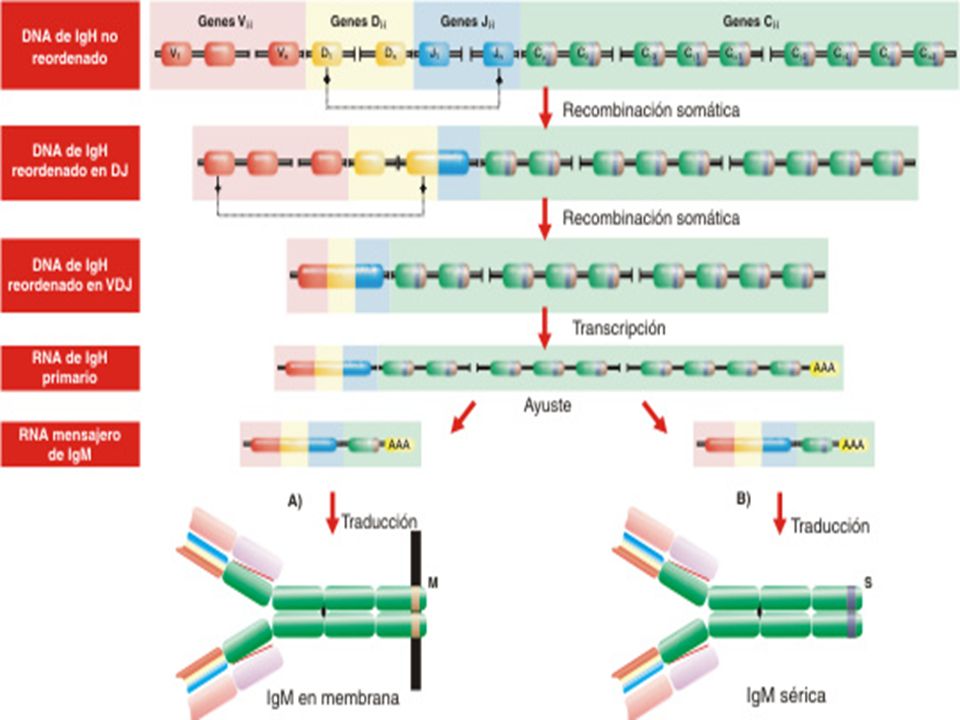
¿ Cómo producen los linfocitos B distintas clases de inmunoglobulinas ?
VDJ Los segmentos que codifican para los dominios constantes de las cadenas pesadas se encuentran hacia el extremo 3’ de los segmentos J. En los humanos la organización es la que se muestra en la parte superior de la figura. Aunque hay pequeñas diferencias para cada gen, en general, existen una serie de exones para cada dominio de la región constante, y un exon para la región bisagra si la clase en cuestión la posee. Además de estos en el extremo 3’ puede utilizarse un exón correspondiente a la forma secretadad (SC), como ocurre en los plasmocitos; o bien una región transmembrana (MC) por ejemplo en linfocitos B vírgenes o en células B de memoria. La forma de membrana o secretada se producen a partir de un transcripto primario de RNA común por procesamiento post-transcripcional.
, como ocurre en los plasmocitos; o bien una región transmembrana (MC) por ejemplo en linfocitos B vírgenes o en células B de memoria. La forma de membrana o secretada se producen a partir de un transcripto primario de RNA común por procesamiento post-transcripcional.")
20
Cadena livianas Kappa y Lambda.
Poseen gen CL que codifica los aa, con Kappa 1 segmento Lambda 4 – 6 segmentos. Poseen gen V 3-4 segmentos para genes V, J que codifican los aa de la región variable.

21
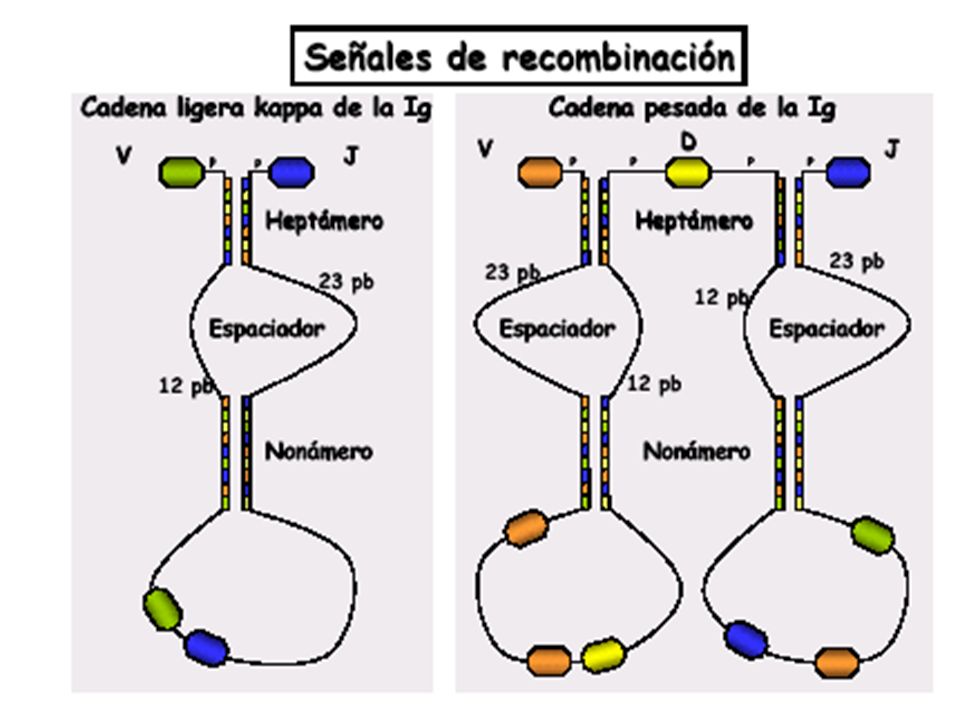
La Recombinación Somática depende de:
La existencia de regiones no conservadas de 23 y 12 pb en los intrones “espaciadores” que separan los segmentos V, D, J y que guían el proceso de recombinación y “secuencias señal” conservadas de 7 pares de bases Heptámeros o 9 pb Nonámeros, reconocidas por las enzimas especializadas. La existencia de recombinasas específicas (RAG1, RAG2) que solo se expresan en los linfocitos B y T inmaduros, con actividad endonucleasa. En los intrones que separan los sementos V, D, y J existen regiones conservadas que guían el proceso de recombinación. Estas regiones están constituidas por heptámeros y nonémeros palindrómicos (opuestos y complentarios). La complementaridad de estas regiones permitela formación del bucle de DNA que se elimina, notar el papel de los nonámeros (de color obscuro en la figura) que al aparearse cierran el bucle. Lo mismo ocurre con los heptámeros, aunque lamentablemente no está claro en la figura. Como las regiones palindrómicas están en el 3’ de los segmentos V y en el 5’ de los J, esto asegura que no se recombinen segmentos V con V, o J con J. Más detalles del proceso se encuentran en el texto de la animación que puede bajarse de (
 que solo se expresan en los linfocitos B y T inmaduros, con actividad endonucleasa. En los intrones que separan los sementos V, D, y J existen regiones conservadas que guían el proceso de recombinación. Estas regiones están constituidas por heptámeros y nonémeros palindrómicos (opuestos y complentarios). La complementaridad de estas regiones permitela formación del bucle de DNA que se elimina, notar el papel de los nonámeros (de color obscuro en la figura) que al aparearse cierran el bucle. Lo mismo ocurre con los heptámeros, aunque lamentablemente no está claro en la figura. Como las regiones palindrómicas están en el 3’ de los segmentos V y en el 5’ de los J, esto asegura que no se recombinen segmentos V con V, o J con J. Más detalles del proceso se encuentran en el texto de la animación que puede bajarse de (")
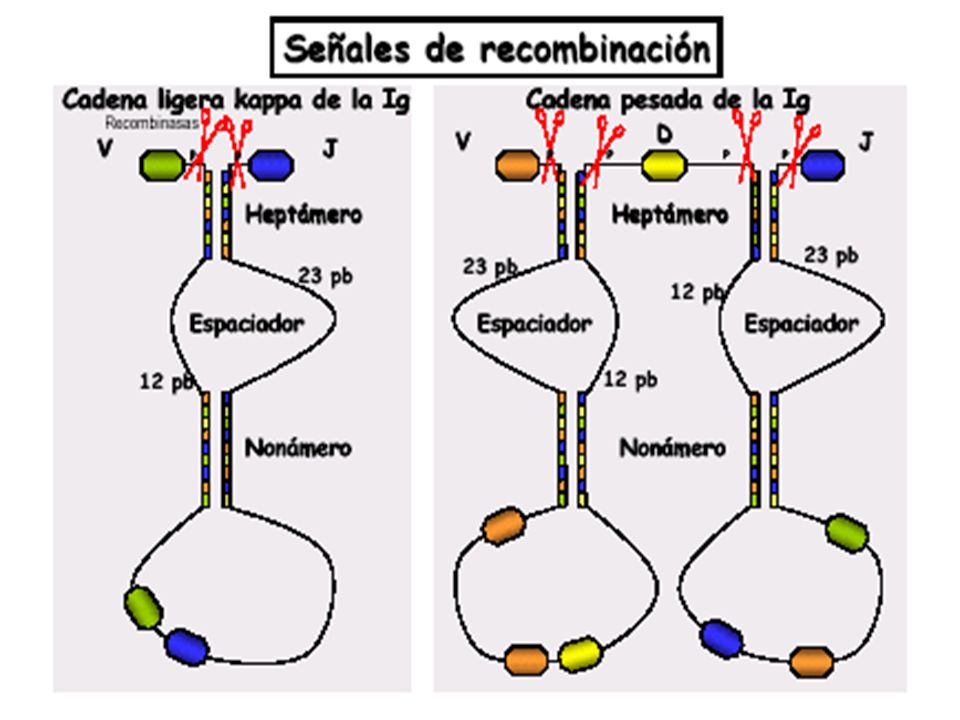
22
Recombinación Formación de estructura intermedia en forma de bucle del DNA , que luego es eliminada en forma de circulo. Extremos sueltos se empalman con la unión de secuencia codificante VJ O JD y cumpliéndose regla 12/23 pb, para el reordenamiento de segmentos flanqueados posteriormente.

28
DIVERSIDAD DE LIGACIÓN O UNIÓN
Durante la union de segmentos V, D, J, se producen variaciones o introducción de codones debido a: La flexibilidad en el corte del bucle terminal que introduce nucleótidos denominados P o palindromicos. b) La acción de la enzima TdT (deoxinucleotidil transferasa terminal) que incorpora nucleotidos (N) en el extremo 3’. Los detalles de este proceso se describen en el texto de la animación incluida en la lámina y que puede bajarse de la página de inmuno (
 La acción de la enzima TdT (deoxinucleotidil transferasa terminal) que incorpora nucleotidos (N) en el extremo 3’. Los detalles de este proceso se describen en el texto de la animación incluida en la lámina y que puede bajarse de la página de inmuno (")
29
La diversidad combinatoria y de ligación concentran la diversificación en el CDR3
Dominio variable CDR1 CDR2 CDR3 N Dominio variable Dominio constante CDR1 CDR2 CDR3 N Ig, cadena pesada (proteína) N V D j gen mRNA () Es muy importante notar que la mayor diversidad combinatoria y de ligación se produce en la región correspondiente al CDR3 tanto para la cadena pesada como liviana. Esto tiene un claro sentido biológico dado que Los aminoácidos que componen los CD3 de ambas cadenas se ubican en la región central del paratope y resultan por tanto cruciales en la interacción con el antígeno.
 N. V. D. j. gen mRNA () Es muy importante notar que la mayor diversidad combinatoria y de ligación se produce en la región correspondiente al CDR3 tanto para la cadena pesada como liviana. Esto tiene un claro sentido biológico dado que Los aminoácidos que componen los CD3 de ambas cadenas se ubican en la región central del paratope y resultan por tanto cruciales en la interacción con el antígeno.")
32
La hipermutación somática es un elemento adicional de generación diversidad
Hipermutación somática : proceso de introducción de mutaciones puntuales en la región variable de los genes de las cadenas livianas y pesadas que expresa el linfocito B. La tasa de mutación es 109 superior a la que ocurre en otras células, y no afecta la región constante de las Igs. Hipermutación somática (órganos linfoides secundarios) Recombinación somática (médula ósea) Hipermutación somática : proceso de introducción de mutaciones puntuales en la región variable de los genes de las cadenas livianas y pesadas que expresa el linfocito B. La tasa de mutación es 109 superior a la que ocurre en otras células, y no afecta la región constante de las Igs. Mientras que los mecanismos de recombianación somática son similares para los linfocitos B y T, la hipermutación somática es exclusiva de las células B.
 Recombinación somática. (médula ósea) Hipermutación somática : proceso de introducción de mutaciones puntuales en la región variable de los genes de las cadenas livianas y pesadas que expresa el linfocito B. La tasa de mutación es 109 superior a la que ocurre en otras células, y no afecta la región constante de las Igs. Mientras que los mecanismos de recombianación somática son similares para los linfocitos B y T, la hipermutación somática es exclusiva de las células B.")
33
Célula dendrítica folicular
La hipermutación somática seguida de la selección por el antígeno, dan lugar al proceso de maduración de la afinidad. Célula B en proliferación Hipermutación somática Centro germinal Progenie con BCRs de distinta afinidad Baja afinidad Alta afinidad Proliferación Célula dendrítica folicular Selección por afinidad La hipermutación somática es el proceso por el cual se introducen mutaciones puntuales en la región variable de los genes de las cadenas livianas y pesadas que expresa el linfocito B, y es de naturaleza aleatoria. La tasa de mutación es 109 superior a la que ocurre en otras células, y no afecta la región constante de las Igs, ni el resto de los genes del linfocito B. Mediante el mecanismo de hipermutación somática, se producen clones mutantes derivados del clon original de linfocito B específico por el antígeno. En estos clones se han introducido en forma aleatoria modificaciones en el paratope que harán que su BCR reconozca con mayor o menor afinidad al antígeno. La posterior selección por el antígeno, rescata los clones de mayor afinidad, y es responsible del fenómeno de MADURACIÓN DE LA AFINIDAD que se observa a medida que transcurre la respuesta inmune (este proceso se verá en detalle más adelante en el curso). En la figura se muestra la respuesta a la inmunización con un hapteno mostrando como a lo largo del proceso se fueron analizando los genes de las cadenas pesadas y livianas de clones de linfocitos B específicos para el hapteno. Nótese que a lo largo de la respuesta, se acumulan mutaciones en las regiones correspondientes a los CDRs, a la vez que se produce un cambio en la cadena pesada (inicialmente IgM pasando a IgG al final). Muerte por apotosis
. En la figura se muestra la respuesta a la inmunización con un hapteno mostrando como a lo largo del proceso se fueron analizando los genes de las cadenas pesadas y livianas de clones de linfocitos B específicos para el hapteno. Nótese que a lo largo de la respuesta, se acumulan mutaciones en las regiones correspondientes a los CDRs, a la vez que se produce un cambio en la cadena pesada (inicialmente IgM pasando a IgG al final). Muerte por apotosis.")
34
El switch de clases permite la secreción de distintos isotipos de Ig a medida que transcurre la respuesta inmune. Debido a que se encuentra situado hacia el 3’ del rearreglo VDJ, el gen de la cadena pesada es el primero en expresarse en el linfocito B inmaduro como receptor de membrana. En la etapa siguiente ocurre la co-expresión de IgM e IgD (contigua a la anterior) en el locus de la cadena pesada de Ig. Esta coexpresión se produce por “splicing” alternativo a nivel del transcripto de RNA, que incluye la regiónes correspondientes a la cadenas pesadas y . La co-expresión de estas cadenas corresponde con un linfocito B maduro, que está listo para salir a la circulación periférica. El encuentro con el antígeno y los eventos que se suceden, particularmente la colaboración con el linfocito T, promueven una nueva recombinación del DNA del linfocito B, y dan lugar al proceso de cambio de clase. Esta recombinación está guiada por secuencias de “switch” presentes en los intrones que separan las regiones correspondientes a las distintas cadenas pesadas. Por estar en los intrones en este caso el rearreglo es siempre productivo. A partir de que se produce un evento de recombinación, no es posible “volver atrás” al isotipo anterior, pero si llevar a cabo un nuevo switch hacia regiones que estén hacia el 3’. El isotipo hacia el cual se produce el cambio de clase está gobernado por los mediadores químicos (citoquínas) que están presentes en el micro-ambiente donde se encuentra el linfocito B, y que son secretados por el linfocito T colaborador.
 en el locus de la cadena pesada de Ig. Esta coexpresión se produce por splicing alternativo a nivel del transcripto de RNA, que incluye la regiónes correspondientes a la cadenas pesadas y . La co-expresión de estas cadenas corresponde con un linfocito B maduro, que está listo para salir a la circulación periférica. El encuentro con el antígeno y los eventos que se suceden, particularmente la colaboración con el linfocito T, promueven una nueva recombinación del DNA del linfocito B, y dan lugar al proceso de cambio de clase. Esta recombinación está guiada por secuencias de switch presentes en los intrones que separan las regiones correspondientes a las distintas cadenas pesadas. Por estar en los intrones en este caso el rearreglo es siempre productivo. A partir de que se produce un evento de recombinación, no es posible volver atrás al isotipo anterior, pero si llevar a cabo un nuevo switch hacia regiones que estén hacia el 3’. El isotipo hacia el cual se produce el cambio de clase está gobernado por los mediadores químicos (citoquínas) que están presentes en el micro-ambiente donde se encuentra el linfocito B, y que son secretados por el linfocito T colaborador.")
35
RESUMEN: Formas en que se genera la diversidad en el dominio variable de las Inmunoglobulinas.
-Diversidad de la línea germinal -Recombinación somática Diversidad combinatoria (rearreglo V,D,J) Diversidad de ligación Combinación de cadenas livianas y pesadas Diversidad potencial = Hipermutación somática El funcionamiento del sistema inmune adaptativo requiere contar con un repertorio muy amplio de receptores para antígenos, de modo de contar con la especificidad adecuada para reconocer los antígenos presentes en los patógenos. La existencia de un importante número de segmentos V, D, J, heredados (diversidad de la línea germinal), junto a los procesos de recombinación e hipermutación somática, producen una enorme diversidad en las moléculas de inmunoglobulinas en los humanos. En el caso del receptor T, la hipermutación somática no tiene lugar, pero los mecanismos anteriores son similares. Existen un número similar de segmentos V, menor número de segmentos D, pero mayor desegmentos J. La diversidad de ligación produce en este caso una mayor diversidad, y además los segmentos D de los TCR tienen todos los marcos de lectura abiertos. Aunque otras especies como el ratón tienen mecanismos muy similares, otros vertebrados difieren drásticamente en la forma que generan la diversidad de sus receptores. En las aves o ovejas hay poca o ninguna diversidad de la línea germinal, y el rearreglo de los segmentos VDJ produce Ig idénticas o similares. En estos casos mecanismos adicionales de conversión de genes (aves) o intensos procesos de hipermutación somática son el origen de la diversificación.. Receptor T: Mecanismos similares operan en el caso del TCR, con excepción de la hipermutación somática que no existe, y la falta de exclusión alélica en el rearreglo de la cadena alfa.
 Diversidad de ligación Combinación de cadenas livianas y pesadas Diversidad potencial = Hipermutación somática. El funcionamiento del sistema inmune adaptativo requiere contar con un repertorio muy amplio de receptores para antígenos, de modo de contar con la especificidad adecuada para reconocer los antígenos presentes en los patógenos. La existencia de un importante número de segmentos V, D, J, heredados (diversidad de la línea germinal), junto a los procesos de recombinación e hipermutación somática, producen una enorme diversidad en las moléculas de inmunoglobulinas en los humanos. En el caso del receptor T, la hipermutación somática no tiene lugar, pero los mecanismos anteriores son similares. Existen un número similar de segmentos V, menor número de segmentos D, pero mayor desegmentos J. La diversidad de ligación produce en este caso una mayor diversidad, y además los segmentos D de los TCR tienen todos los marcos de lectura abiertos. Aunque otras especies como el ratón tienen mecanismos muy similares, otros vertebrados difieren drásticamente en la forma que generan la diversidad de sus receptores. En las aves o ovejas hay poca o ninguna diversidad de la línea germinal, y el rearreglo de los segmentos VDJ produce Ig idénticas o similares. En estos casos mecanismos adicionales de conversión de genes (aves) o intensos procesos de hipermutación somática son el origen de la diversificación.. Receptor T: Mecanismos similares operan en el caso del TCR, con excepción de la hipermutación somática que no existe, y la falta de exclusión alélica en el rearreglo de la cadena alfa.")
36
TCR

37
RECEPTOR DE LINFOCITOS T (TCR)
Receptor ubicado en la membrana de los clones de los linfocitos T Reconoce antígenos proteicos Se generan a partir de Linfocitos inmaduros del Timo

38
TCR del linfocito T El receptor posee:
una cadena o (simil cadena liviana) una cadena o (simil cadena pesada) Dependiendo si es TCR o TCR
 una cadena o (simil cadena pesada) Dependiendo si es TCR o TCR ")
39
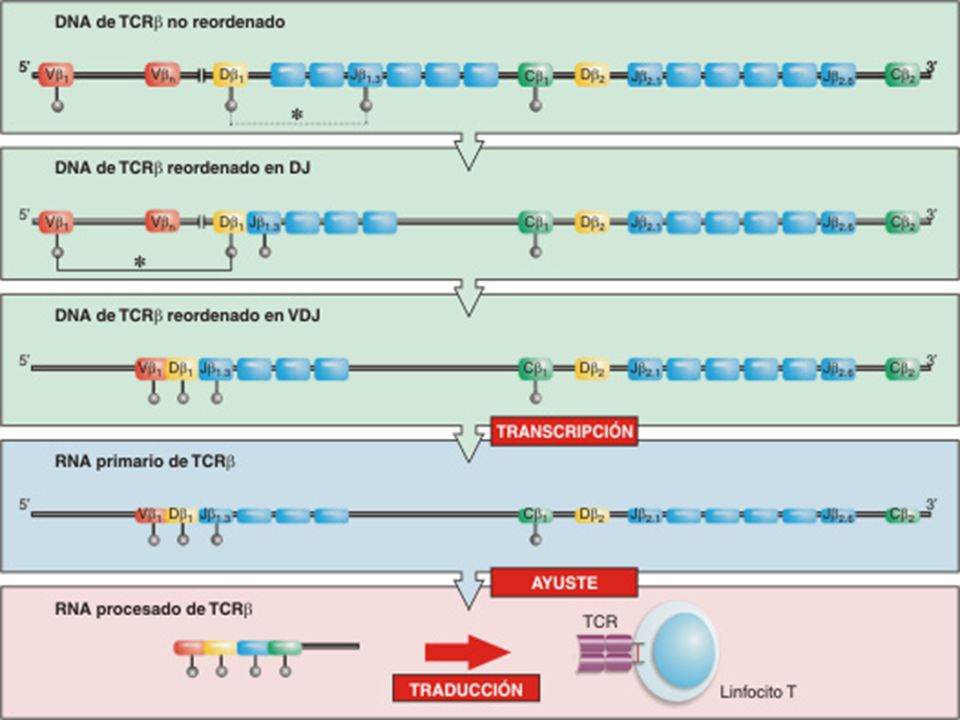
MECANISMO DE RECOMBINACIÓN SOMÁTICA
El reordenamiento de los genes del TCR es un tipo especial de recombinación del ADN que afecta a segmentos génicos no homólogos y está mediado por la actividad coordinada de varias enzimas, algunas de las cuales sólo se encuentran en linfocitos en desarrollo, mientras que otras son enzimas de reparación del ADN.

40
INICIO DE LA RECOMBINACION
Precursor linfático entra en el Timo: timocito que se llaman pro-T, estos no expresan ni CD4 ni CD8 por lo que se llaman dobles negativos. Los dobles negativos empezarán a expresar enzimas para el reordenamiento, recombinasas: RAG (1, 2) y TdT que sólo se expresan en los linfocitos B y T Son esenciales para la srecombinaación Los dobles negativos no tienen receptores.
 y. TdT que sólo se expresan en los linfocitos B y T. Son esenciales para la srecombinaación. Los dobles negativos no tienen receptores.")
44
RECOMBINASAS RAG 1: Ez que une y corta el ADN, es decir, actúa como una endonucleasa específica. Es enzimáticamente activa cuando forma un complejo con el RAG 2 RAG 2: Tiene características estructurales similares al RAG 2 y actúa potenciando a RAG 1 Son enzimas importantes en la recombinación.

45
PROPIEDADES RAG Son específicas de algunos tipos celulares, expresadas sólo en la estirpe linfocítica B y T inmadura. En consecuencia, la recombinasa V(D)J únicamente se activa con linfocitos inmaduros. Se expresan en la fase G0 y G1 del ciclo celular, están silenciados en las células que proliferan 3en el punto2 : no en los maduros, lo que explica porque no continuan produciendose reordenamiento de RCT en los que expresan receptores antigenicos funcionales
J únicamente se activa con. linfocitos inmaduros. Se expresan en la fase G0 y G1 del ciclo celular, están silenciados. en las células que proliferan. 3en el punto2 : no en los maduros, lo que explica porque no continuan produciendose reordenamiento de RCT en los que expresan receptores antigenicos funcionales.")
46
ENZIMAS ACCESORIAS PC- ADN (complejo de la proteína cinasa dependiente de ADN): Su función es la reparación de las roturas en la doble cadena de ADN provocadas por la recombinasa Se expresan en muchos tipos celulares

47
DIVERSIDAD Es producto de:
Diversidad de combinación producto de la suma de las combinatorias posibles de las cadenas asociadas. “Recombinación Somática” Diversidad de las uniones en el momento del empalme de exones adición(TdT) o perdida de nucleótidos(endonuleasas)
 o perdida de nucleótidos(endonuleasas)")
Presentaciones similares