Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Citoesqueleto y movilidad celular
M. en C. Carlos Gerardo Castillo Sosa FMVZ-BUAP Primavera 2012

2
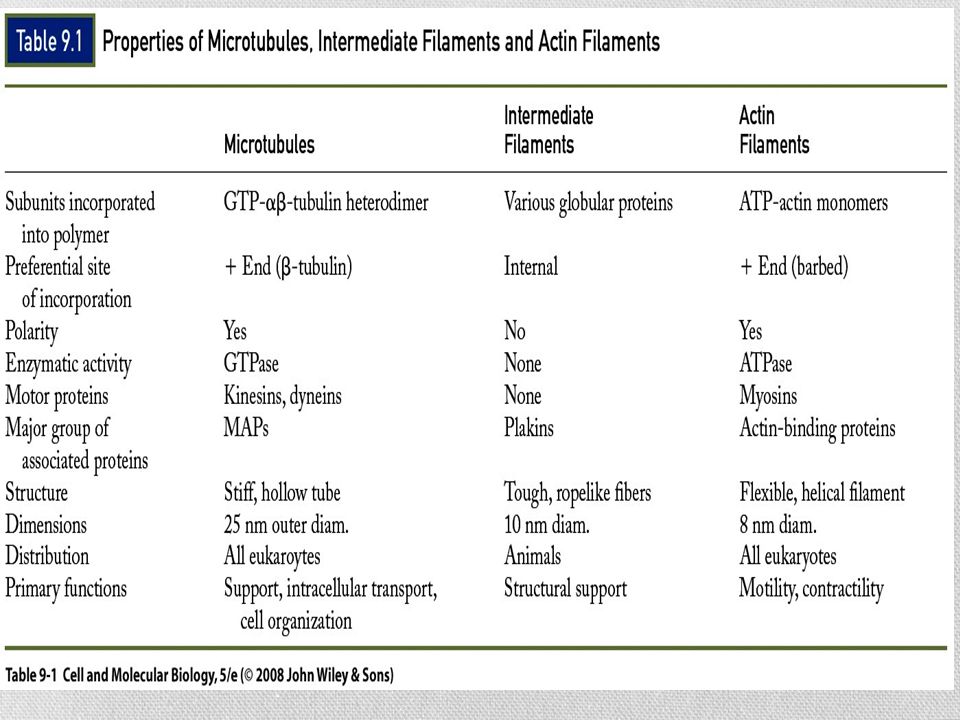
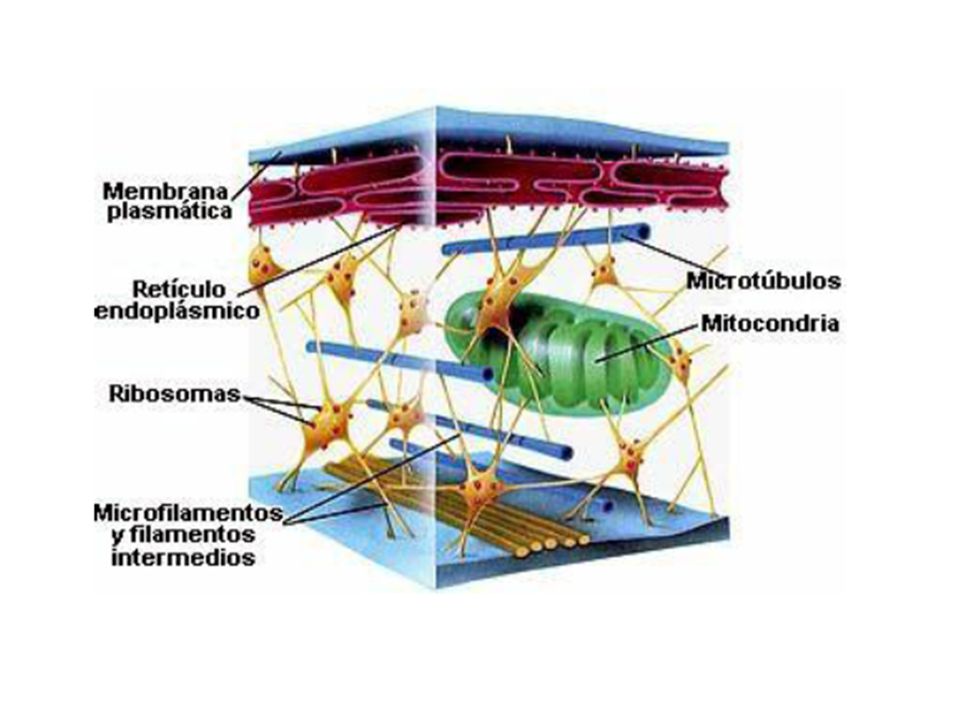
El Citoesqueleto Compuesto de tres estructuras filamentosas bien definidas: Microtúbulos Microfilamentos Filamentos intermedios Son polímeros de subunidades proteicas unidas por enlaces no covalentes

4
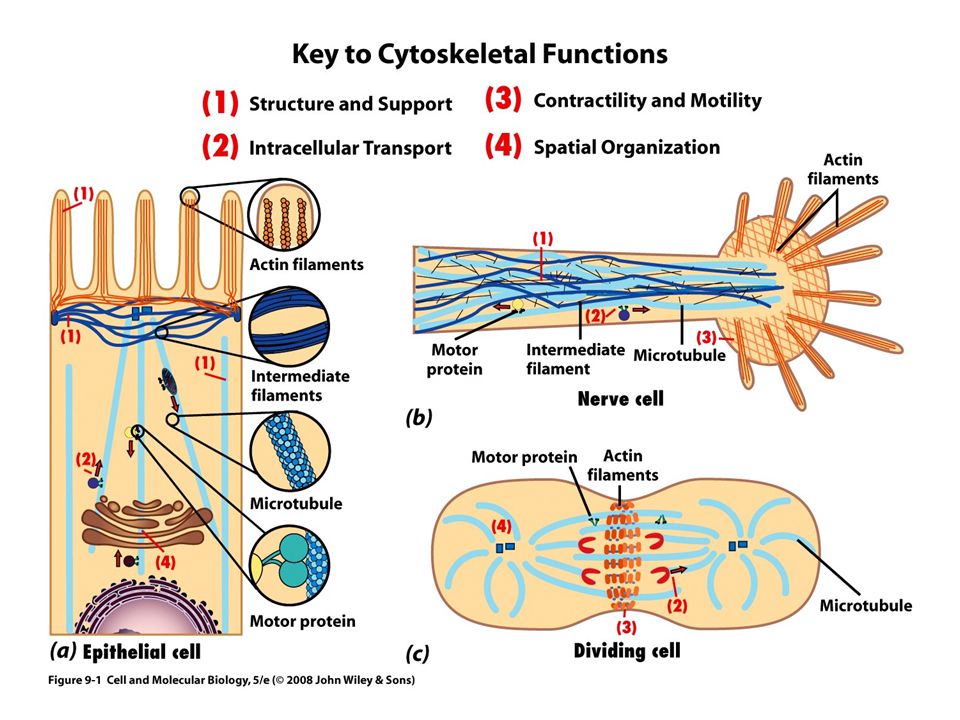
Revisión de las funciones del citoesqueleto
Un andamio dinámico que brinda soporte estructural Un marco interno que establece las posiciones de los organelos Una red de rieles El aparato generador de fuerza que mueve la célula de un sitio a otro Componente esencial para la división celular

9
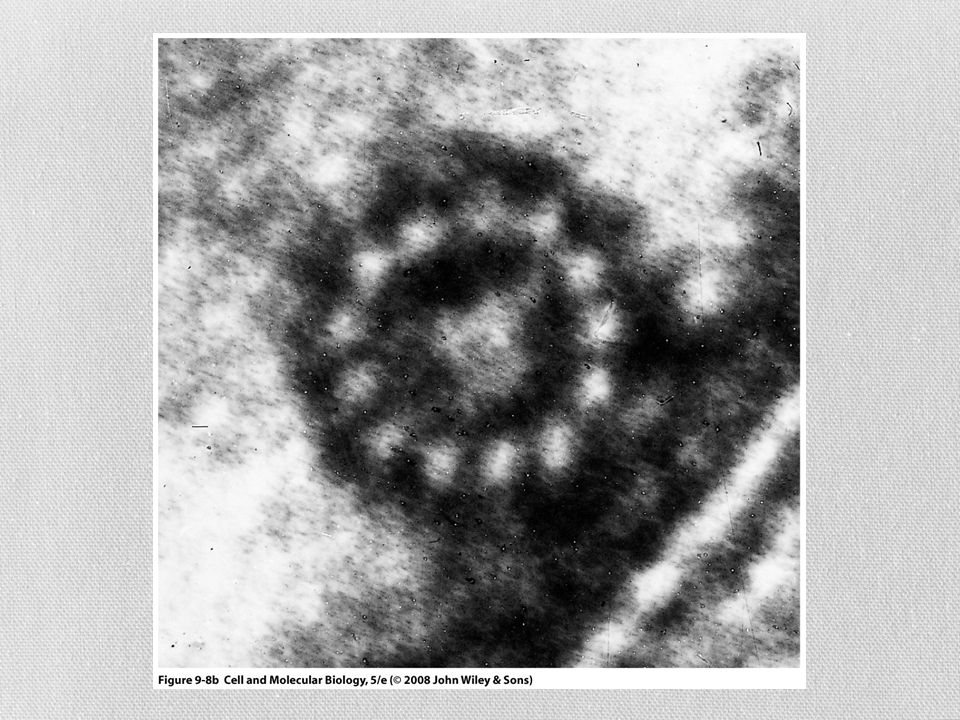
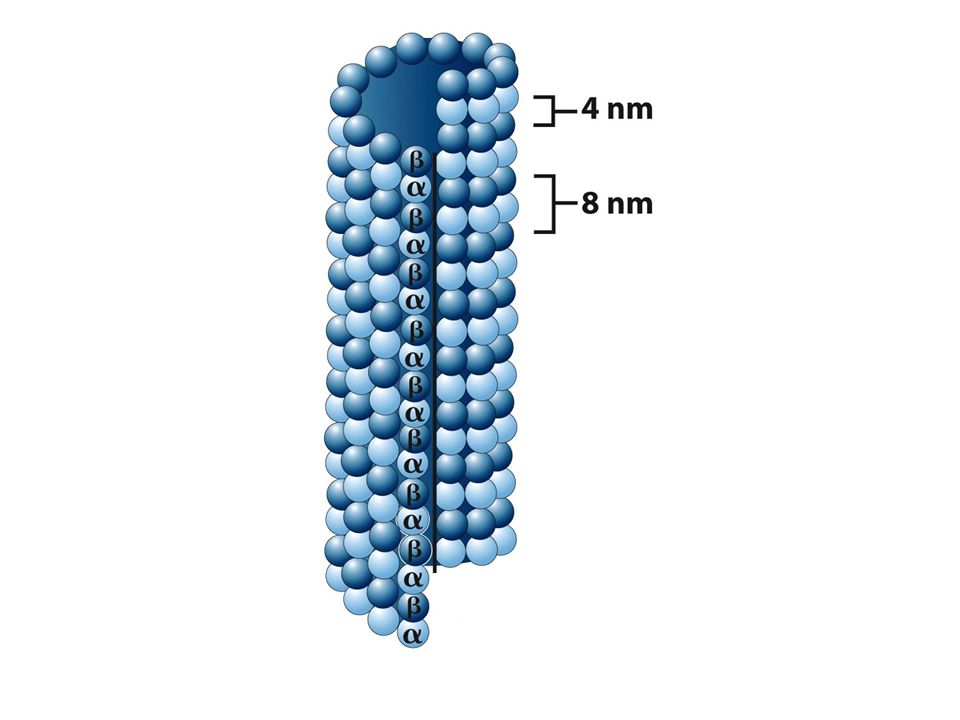
Microtúbulos Son estructuras tubulares huecas
Forman huso mitótico y centro de cilios y flagelos La pared esta constituida por hileras longitudinales llamadas protofilamentos Cada protofilamento esta ensamblado por bloques dimericos Tubulina α y tubulina β

10
Microtúbulos Un extremo del microtúbulo termina con una fila de subunidades beta y se llama extremo mas El extremo contrario termina con subunidades alfa y se llama extremo menos

13
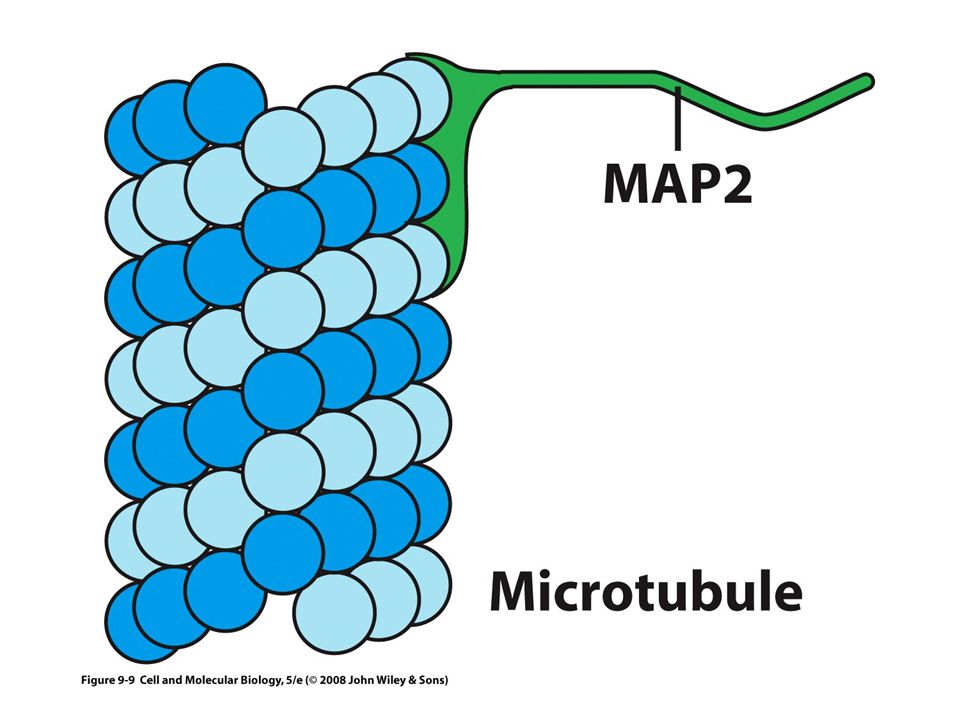
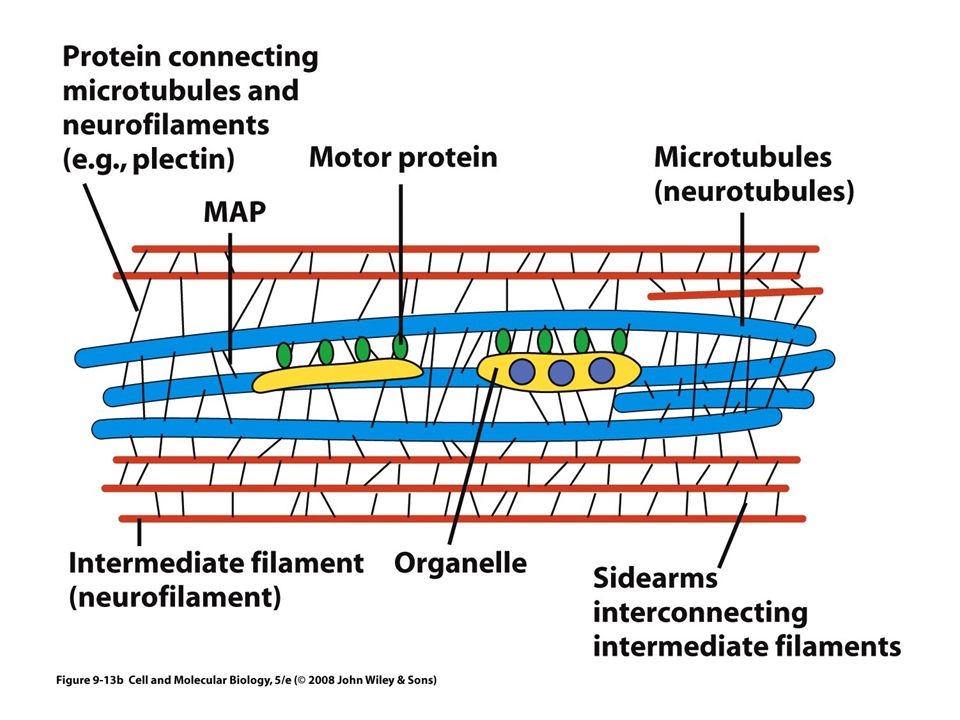
Las MAP Proteínas asociadas a microtúbulos ( Microtubules Asociated Proteins) Incrementan la estabilidad de los microtúbulos y promueven su ensamble La actividad de las MAP se controla principalmente con la adición y el retiro de grupos fosfato

15
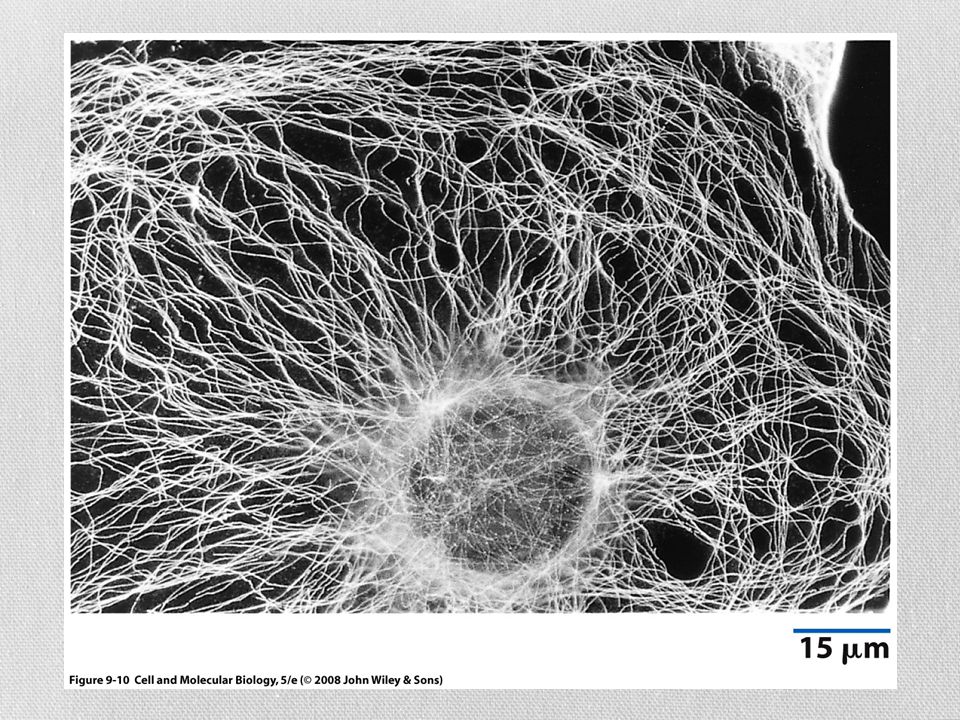
Microtúbulos como soportes y organizadores estructurales
La distribución de los microtúbulos ayuda a determinar la forma de la célula También participan en el mantenimiento de la organización interna de la célula

17
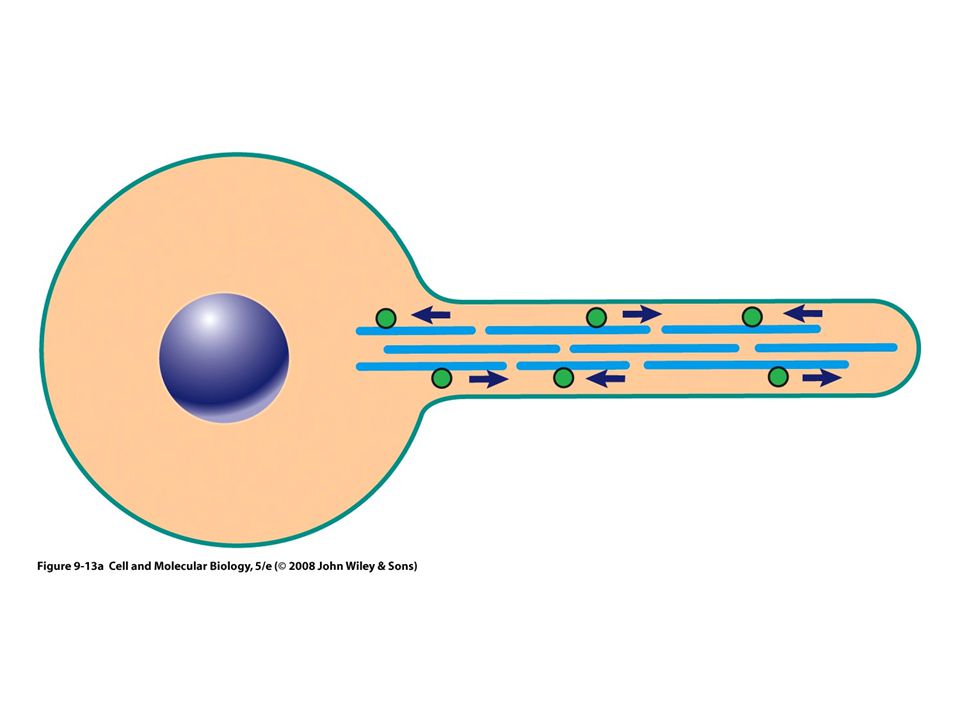
Microtúbulos como agentes de motilidad intracelular
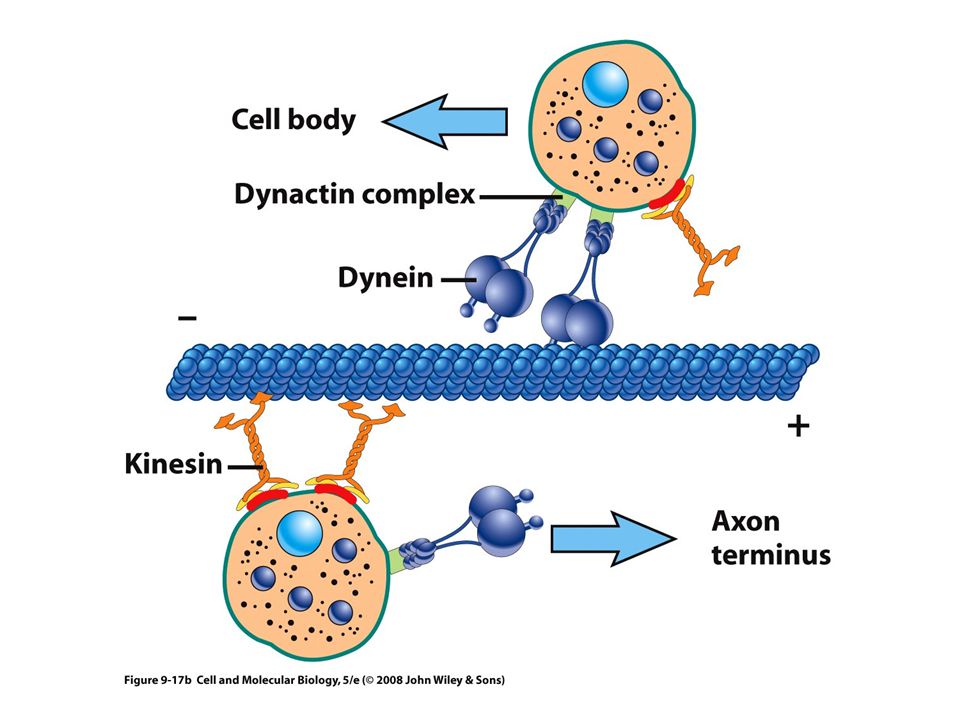
Transporte axónico La mayor parte de los materiales de una neurona motora, como moléculas neurotransmisoras, se incluyen en compartimentos dentro de vesículas membranosas Luego se transportan por toda la longitud del axón 0.5 µm por segundo Dirección anterógrada

20
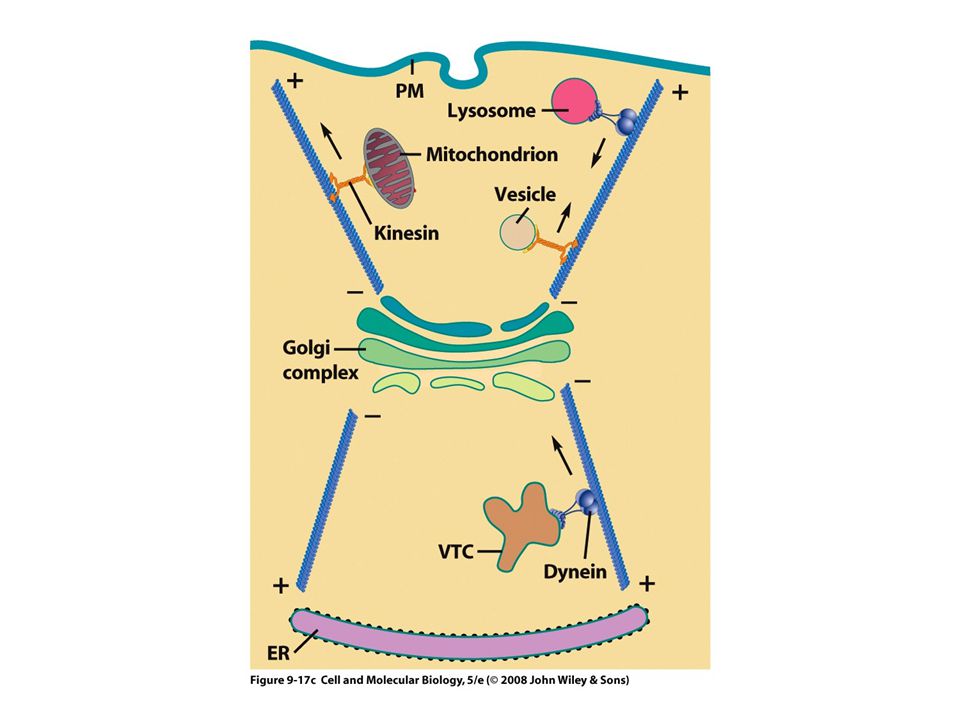
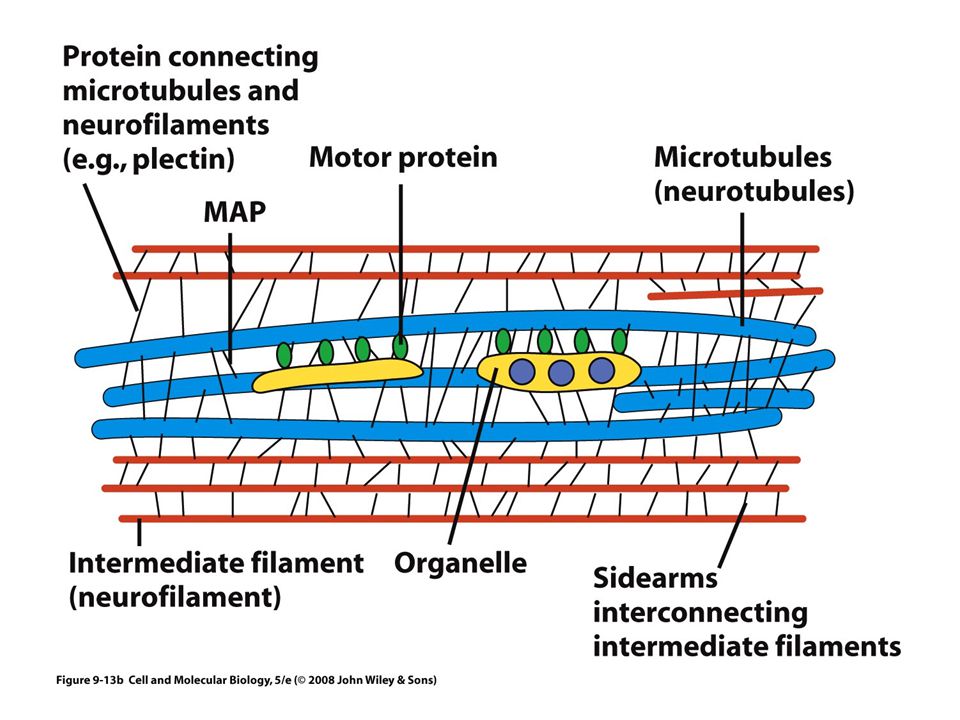
Proteínas motoras Estas proteínas convierten la energía química (ATP) en energía mecánica Transportan: vesículas, mitocondrias, lisosomas y otros filamentos del citoesqueleto Tres grandes familias: Cinesinas Dineínas Miosinas Los filamentos intermedios no tienen motores proteicos

21
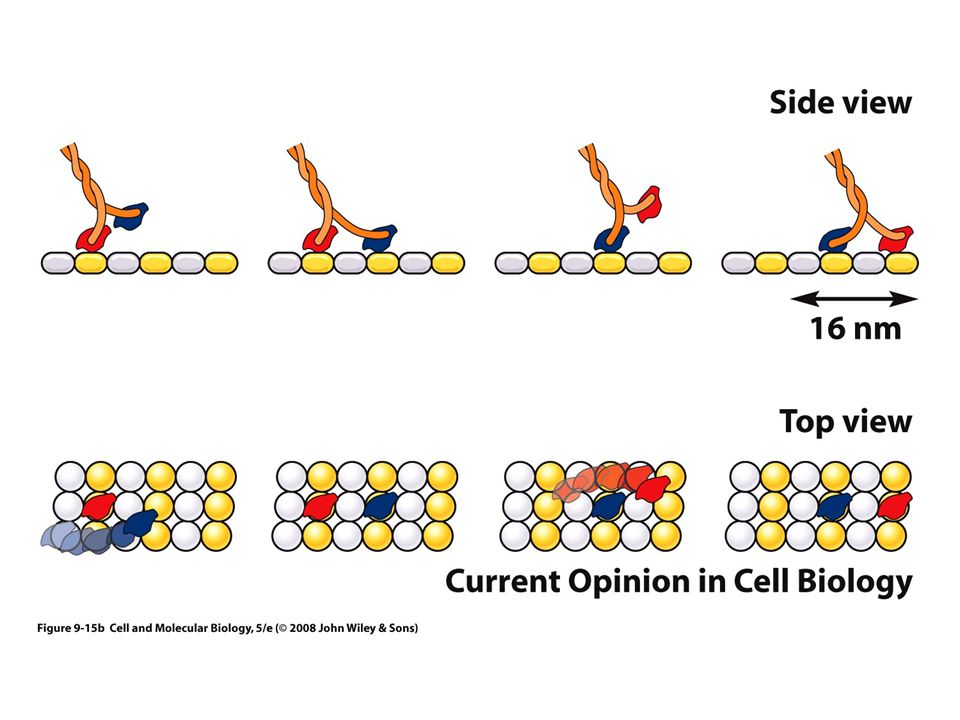
Las proteínas motoras se mueven por pasos en una sola dirección a lo largo del riel de citoesqueleto
Sufre un cambio de conformación: ciclo mecánico Ciclo químico: Unión de una molécula de ATP con el motor, Hidrólisis del ATP Unión a una nueva molécula de ATP Proporciona un “golpe” que mueve el motor

22
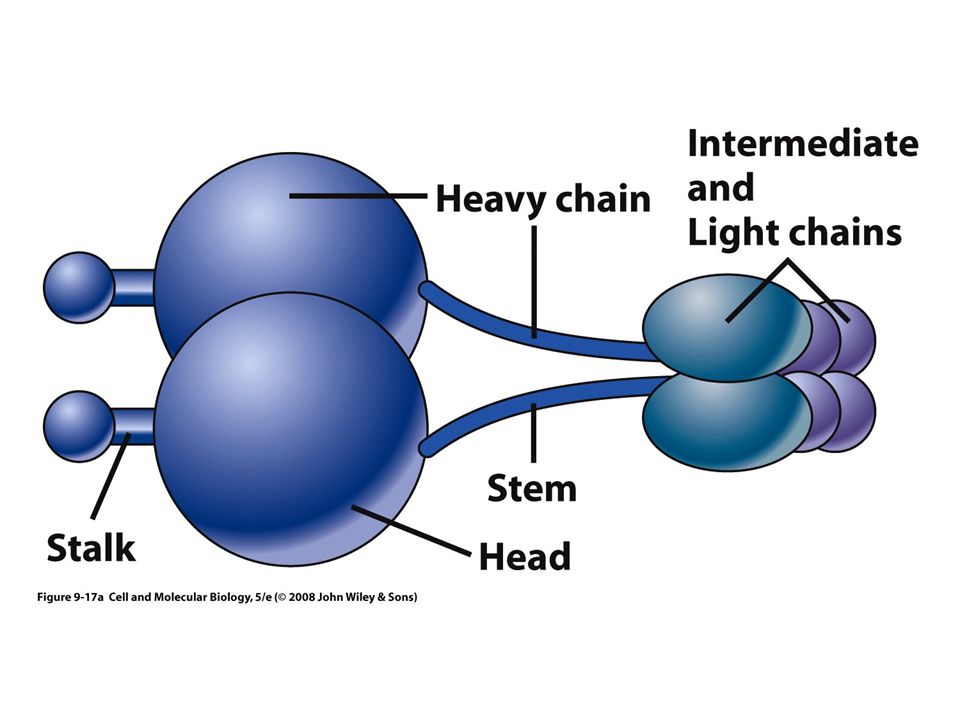
Cinesinas Las proteínas motoras mejor estudiadas
Tetramero, dos cadenas ligeras y dos pesadas Tienen un par de cabezas globulares que se unen al microtúbulo y actúan como “maquinas” generadoras de fuerza Hidrolizan ATP Cabeza, cuello y cola con forma de abanico

23
Cadena pesada Cadena ligera Bisagra flexible Cabezas Cuello Tallo Cola

25
Una sola molécula de cinesina se mueve por un solo protofilamento de un microtúbulo
Velocidad proporcional a la concentración de ATP 0.1 µm por segundo El movimiento es muy progresivo Superfamilia de proteínas conocidas como KLP Los mamíferos producen hasta 45 KLP distintas

26
Las porciones motoras de las KLP tienen secuencias de aminoácidos relacionadas
Las colas de las KLP tienen secuencias diversas La cinesina y la mayoría de las KPL se mueven hacia el extremo mas del microtúbulo

27
Dineinas Es una proteína muy grande, formada por dos cadenas pesadas idénticas y varias cadenas intermedias y ligeras Cada cadena pesada consiste en una cabeza globular grande con dos proyecciones alargadas (tallo) Actúa como una maquina generadora de fuerza Tiene una proyección larga, conocida como pie, que esta relacionada con el cargamento
 Actúa como una maquina generadora de fuerza. Tiene una proyección larga, conocida como pie, que esta relacionada con el cargamento.")
29
La dineína se mueve hacia el extremo menos del microtúbulo
Dos funciones principales: Un agente generador de fuerza para el posicionamiento del huso y el movimiento de los cromosomas durante la mitosis Un motor microtubular dirigido al extremo menos para el movimiento de organelos y vesículas

32
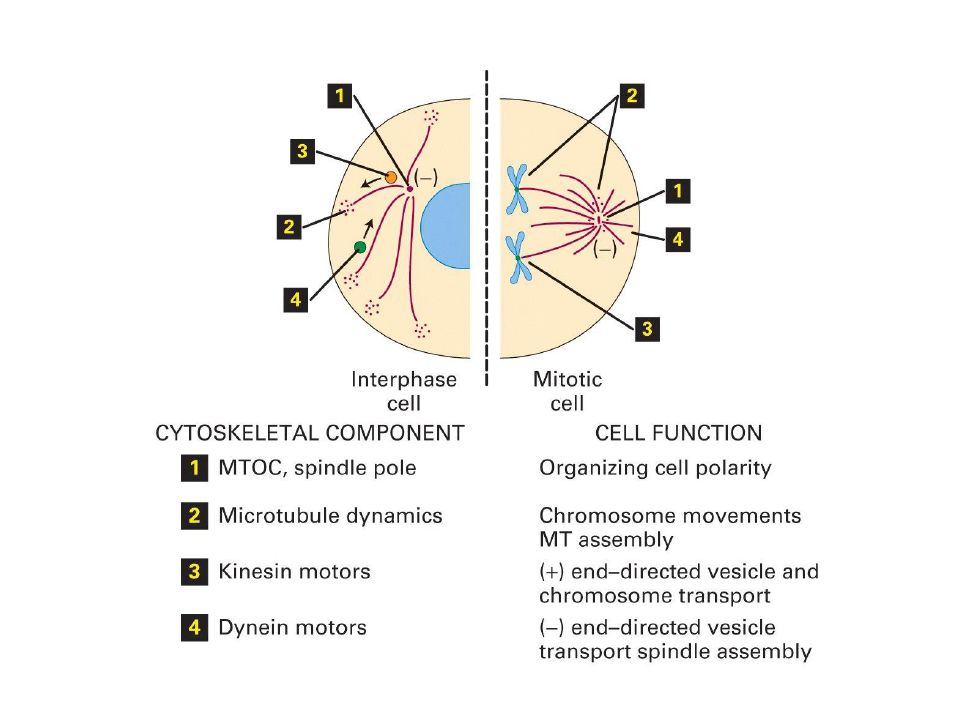
Centros organizadores de los microtúbulos
La función de un microtúbulo en la célula dependerá de su localización Ensamble de microtúbulos en dos fases: Nucleación, fase lenta Elongación, fase rápida Relacionadas con los centros organizadores de microtúbulos (MTOC)
")
34
Los MTOC tienen funciones similares en todas las células
Controlan el numero de microtúbulos Su polaridad Numero de protofilamentos Momento y localización de su ensamble Tubulina gamma

35
Función de la tubulina gamma en el
ensamblaje de los microtúbulos

36
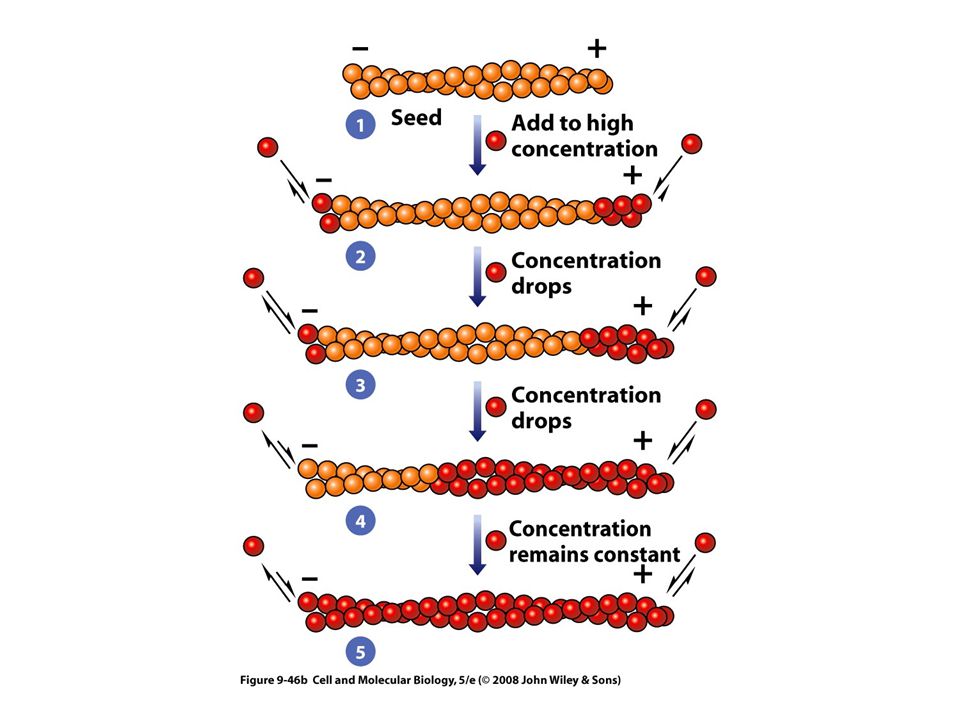
Las propiedades dinámicas de los microtúbulos
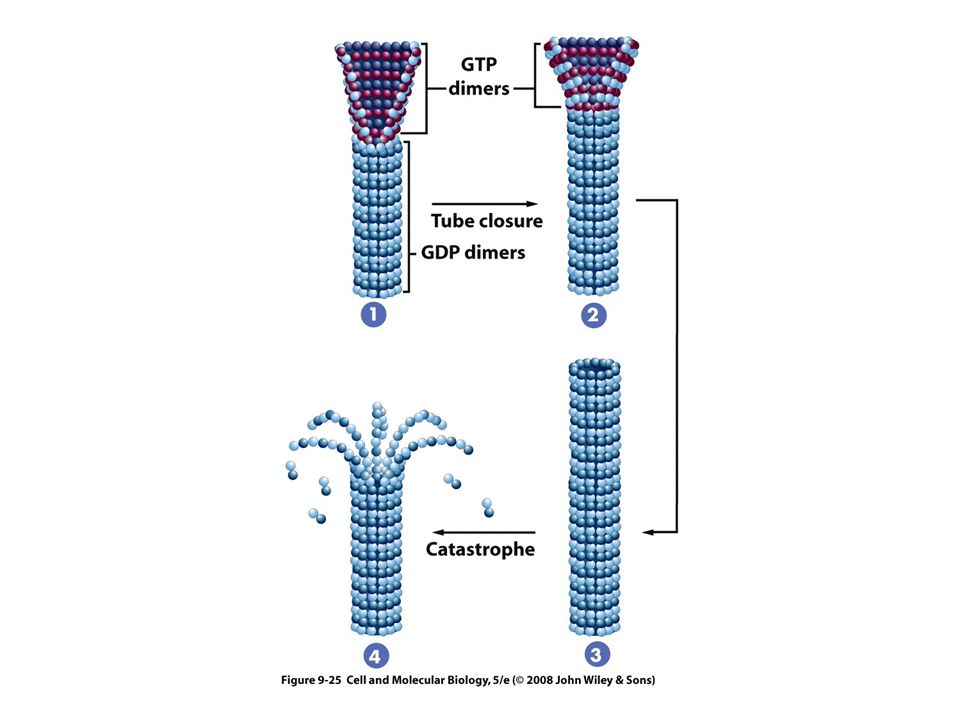
Presentan diferencias en su estabilidad Los microtúbulos del huso mitótico son muy lábiles Los microtúbulos de las neuronas son mucho menos lábiles Los de cilios y flagelos son muy estables La labilidad refleja el hecho de que los microtúbulos son polímeros formados por enlaces no covalentes

37
Los microtúbulos del citoesqueleto están sujetos a despolimerización y repolimerizacion
Para el ensamble se necesita GTP El GTP se une con la tubulina beta La incorporación del dímero no necesita de la hidrólisis de GTP El GTP se hidroliza poco después y el GDP resultante permanece unido al polímero permanentemente

38
Disposiciones de los microtúbulos durante el ciclo celular de una
célula vegetal

41
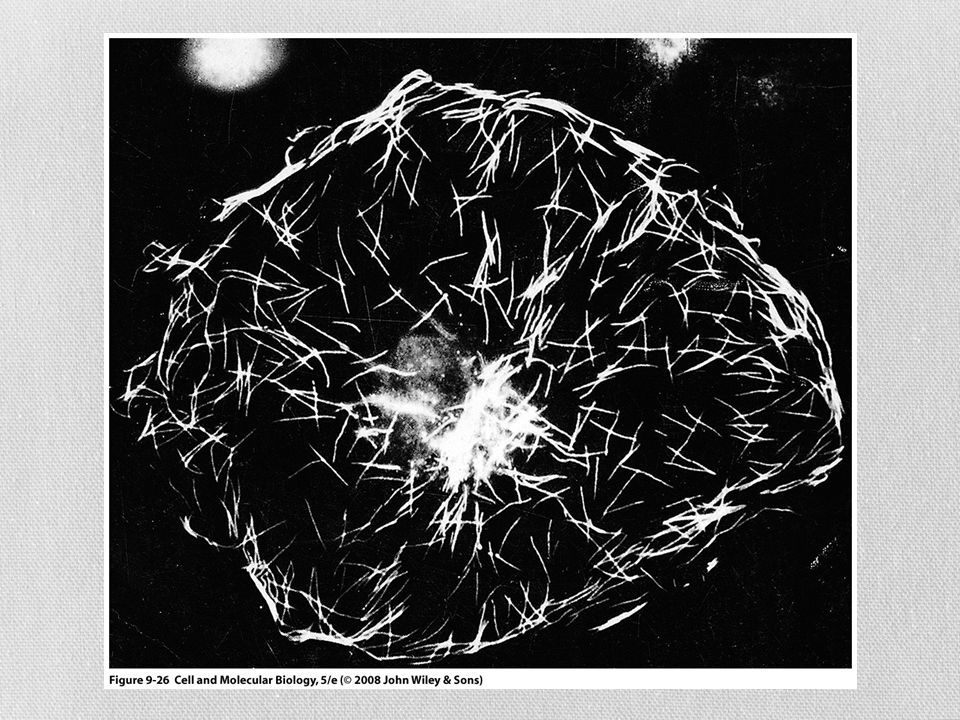
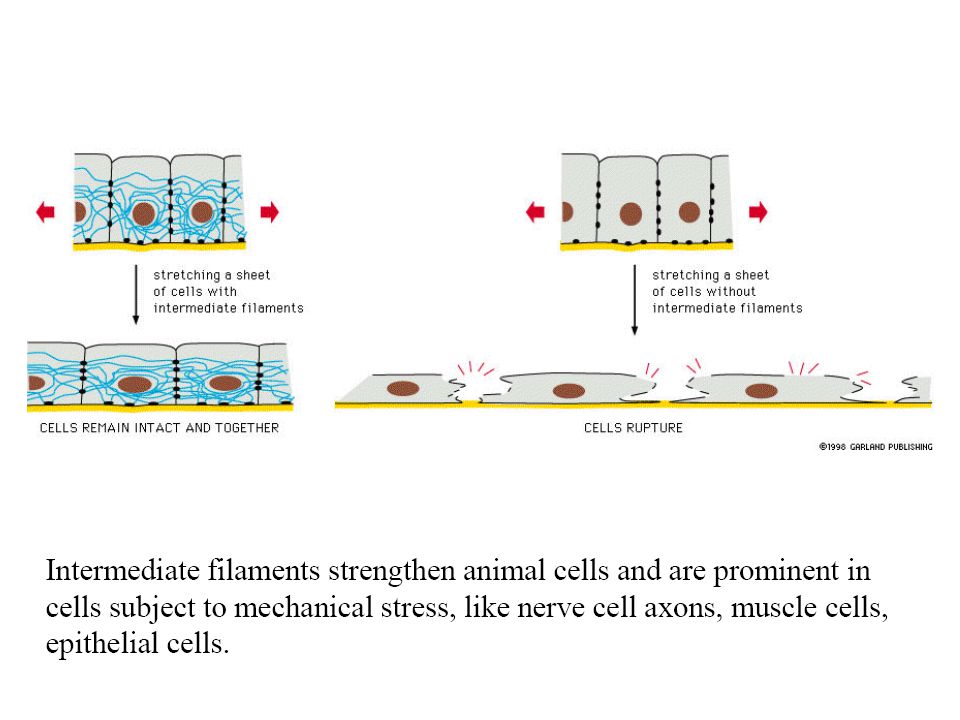
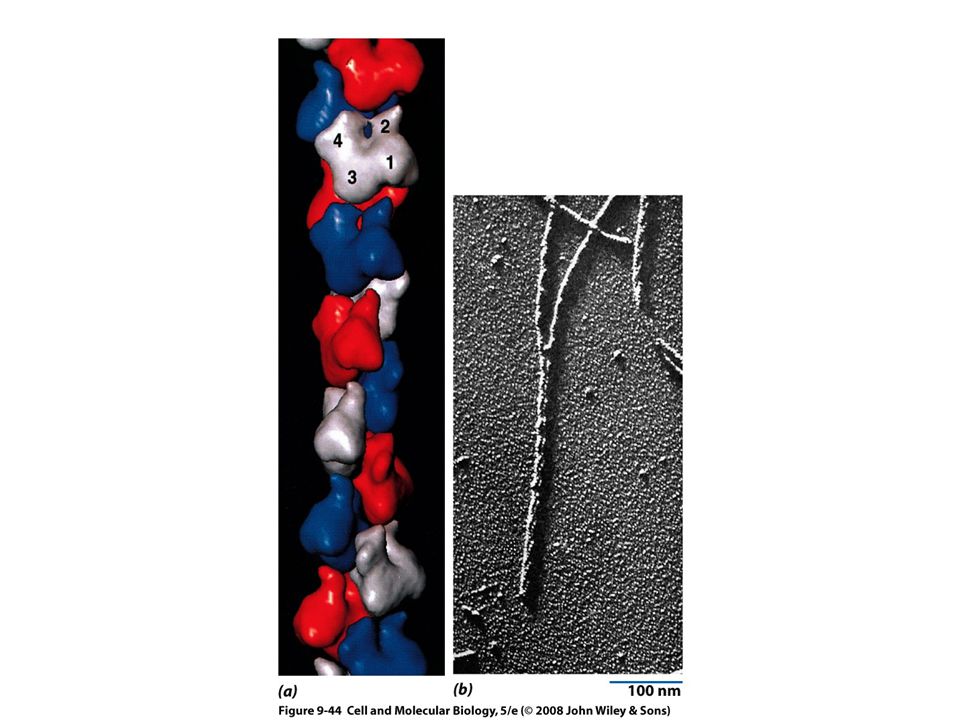
Filamentos intermedios
Son fibras fuertes, similares a cuerdas Proporcionan fuerza mecánica a las células que se someten a tensión física Neuronas Células musculares Células epiteliales de cavidades

42
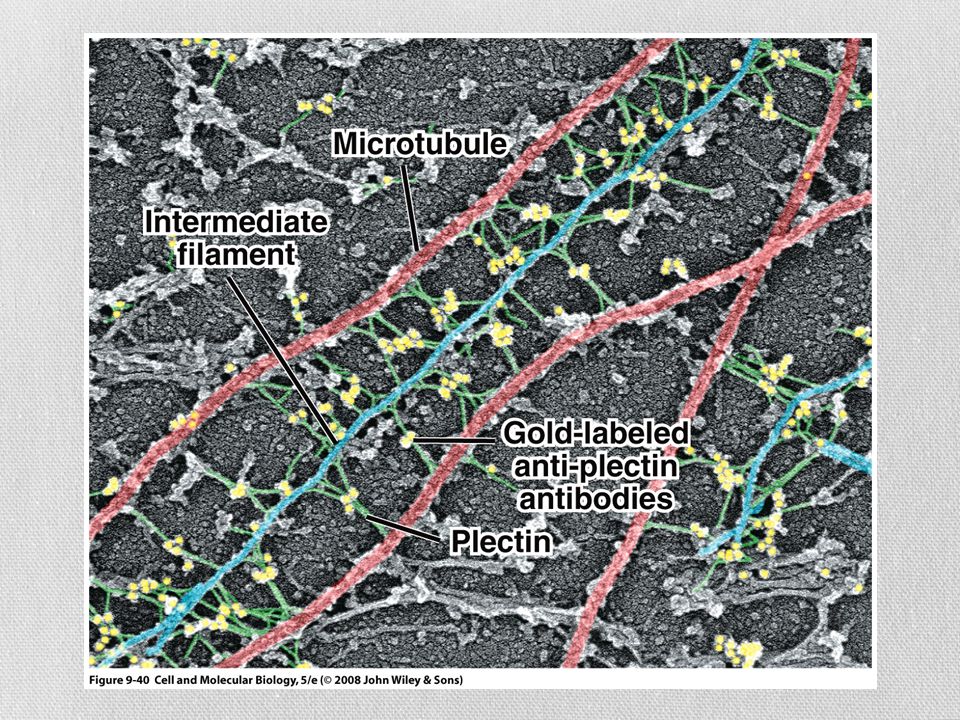
Filamentos intermedios
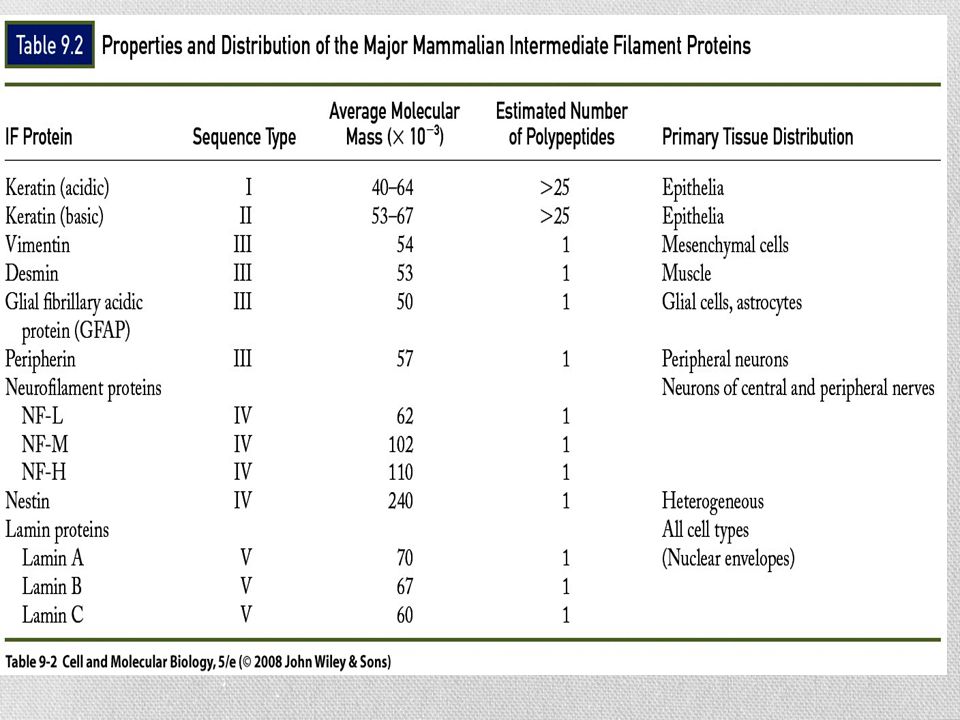
Se han identificado solo en células animales Se conectan con otros tipos de filamentos por medio de una proteína llamada plectina Estructuralmente heterogéneo Cinco clases principales de subunidades De acuerdo a su distribución tisular

46
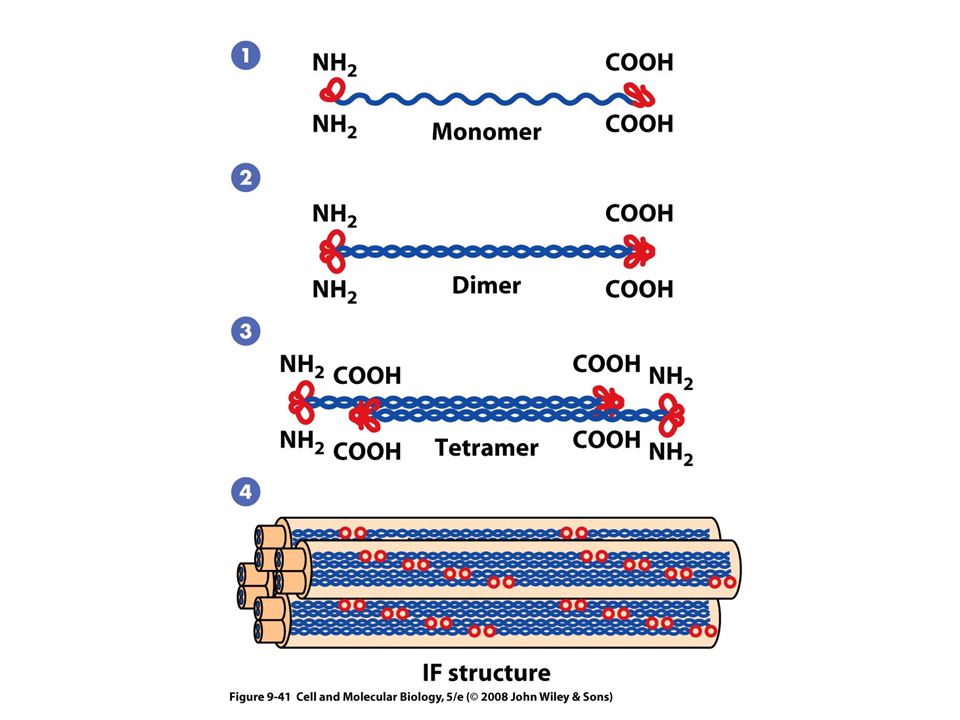
Ensamble y desensamble de los filamentos intermedios
Unidad básica formada por un tetrámero El filamento ensamblado carece de polaridad Menos sensibles a otros agentes químicos Mas difíciles de disolver Tienen un comportamiento dinámico in vivo

48
Tipos y funciones de los filamentos intermedios
Los filamentos de queratina constituyen las principales proteínas estructurales de las células epiteliales Hepatocitos Células epidérmicas Células acinares pancreáticas En neuronas, neurofilamentos Formados por 3 proteínas distintas: NF-L, NF-H y NF-M

50
Ratones modificados genéticamente
Delecion en el gen que codifica para K14 Polipéptido de queratina, encontrado en células de epidermis Muy sensibles al mas mínimo traumatismo Epidermolisis ampollar simple Miopatía relacionada con desmina Enfermedad de Lou Gehrig o esclerosis lateral amiotrofica

52
Microfilamentos Miden alrededor de 8 nm de diámetro
Compuesto de subunidades globulares de una proteína llamada actina Un filamento de actina tiene dos estructuras helicoidales Cada filamento tiene un extremo + y otro – Según el tipo de célula y la actividad, los filamentos pueden organizarse en conjuntos ordenados, en redes laxas o en haces anclados con firmeza

54
Microfilamentos La actina se reconoce como una proteína principal en todos los tipos de células eucariotas Las especies animales y vegetales tienen varios genes codificadores de actina Las actinas se han conservado de manera notable a lo largo de la evolución Moléculas de actina de distintas fuentes pueden polimerizarse juntas

55
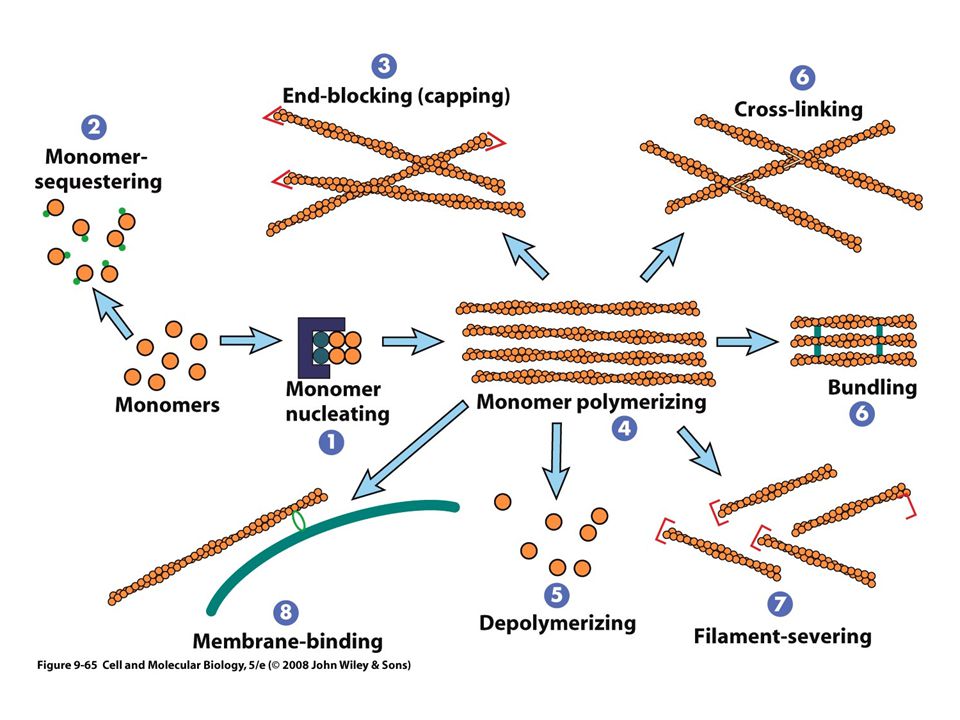
Ensamble y desensamble de Microfilamentos
Un monómero de actina debe unirse a un ATP La actina funciona como una ATP-asa La función del ATP, es similar a la de GTP El ATP unido al monómero de actina se hidroliza después de su incorporación al filamento de actina El filamento consiste en subunidades ADP- actina

57
Las células mantienen un equilibrio dinámico entre las formas monomericas y polimericas de la actina
El ensamble y desensamble de los filamentos dependerá de varias proteínas “accesorias” Cambios locales en una región particular de la célula La reorganización ese necesaria para los procesos como locomoción celular

58
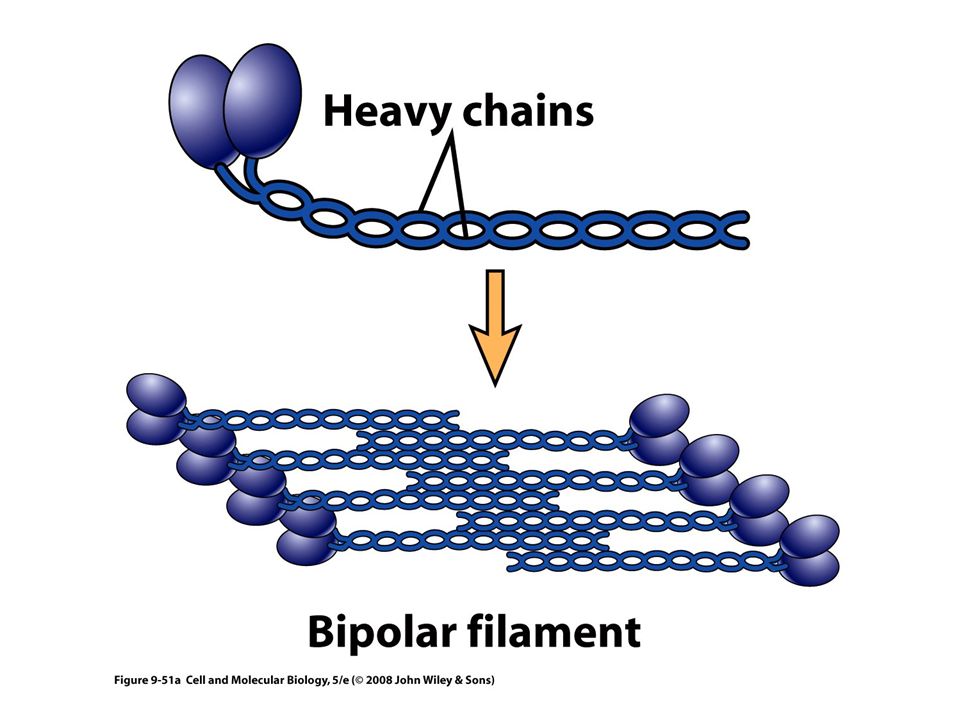
Miosina Todos los motores conocidos que funcionan con actina pertenecen a la superfamilia de la miosina En general la miosina se mueve hacia el extremo mas del filamento La miosina se encuentra en: Tejido muscular esquelético Tejido muscular liso Tejido muscular cardiaco Tejido extramuscular

59
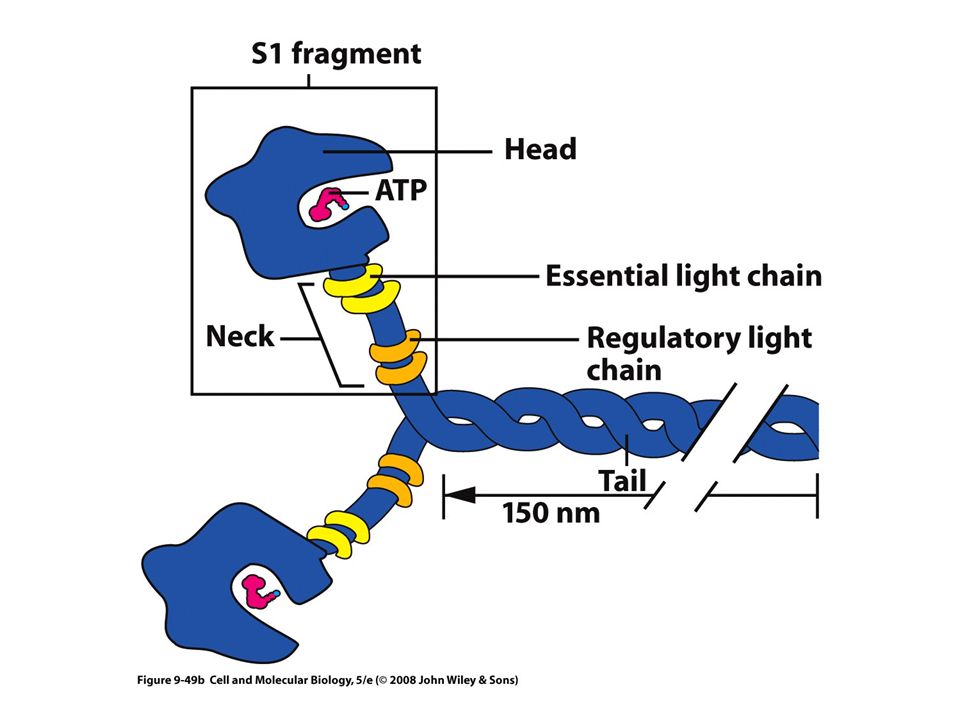
Miosina Tienen un dominio motor característico (cabeza)
La cabeza tiene un sitio para unirse a la actina y otro para el ATP Los dominios de cabeza son similares, los de la cola son muy diversos Se dividen en dos grupos: Miosinas convencionales o Tipo II Miosinas no convencionales Tipos I y III al XVIII

62
Miosinas no convencionales
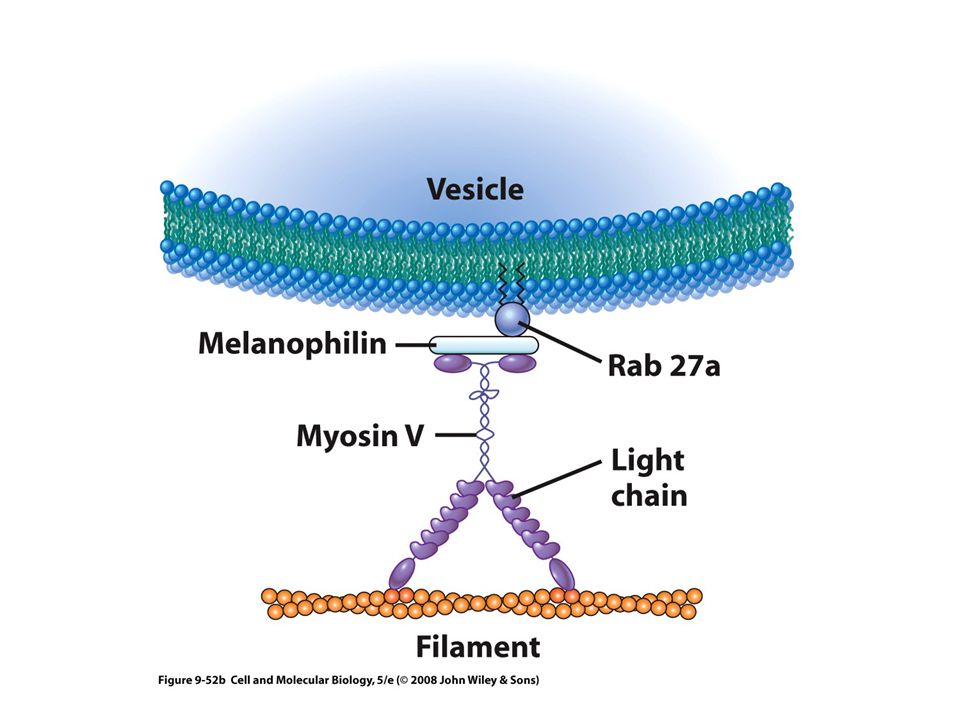
Varias se relacionan con diversos tipos de vesículas citoplasmáticas (miosinas I, V y VI) Algunas vesículas contienen motores basados en microtúbulos y motores basados en Microfilamentos Cooperación entre microtúbulos y microfilamentos estudiada en células pigmentarias Transporte de melanosomas Síndrome de Griscelli
 Algunas vesículas contienen motores basados en microtúbulos y motores basados en Microfilamentos. Cooperación entre microtúbulos y microfilamentos estudiada en células pigmentarias. Transporte de melanosomas. Síndrome de Griscelli.")
63
Interacciones entre la miosina y la cinesina

65
Zona de oclusión

66
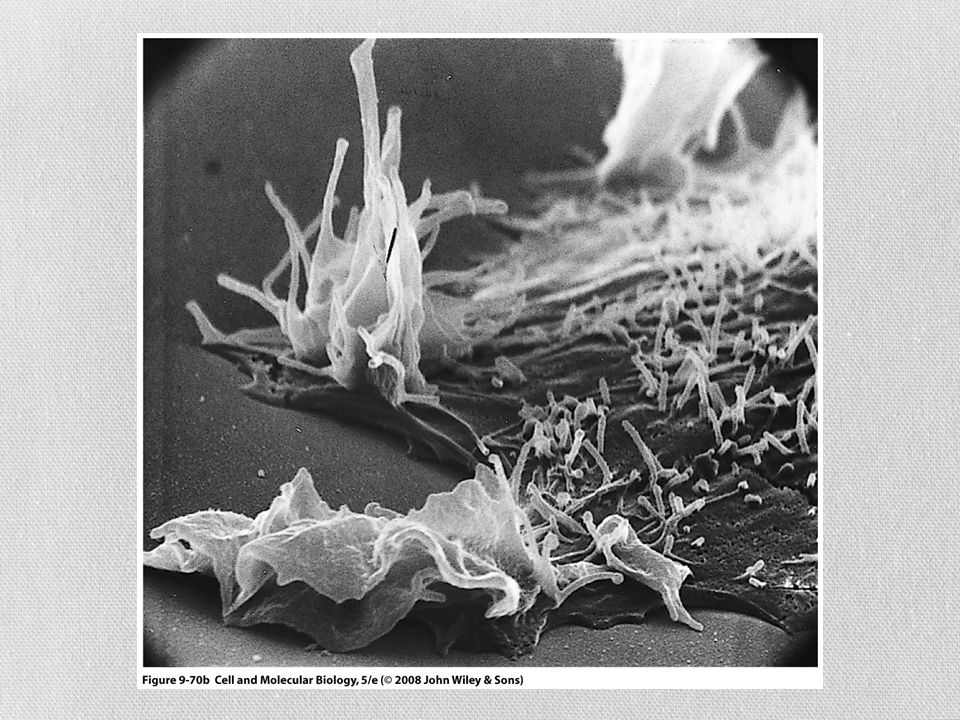
Micrografía electrónica de barrido de células vellosas de la cóclea de ratón

67
Proteínas de unión a actina
La organización y el comportamiento de los filamentos de actina dependen de las proteínas de unión Afecta el ensamble y desensamble, sus propiedades físicas y su interacción Las proteínas de unión a actina pueden dividirse en varias categorías según su función en la célula

68
Proteínas de unión a actina
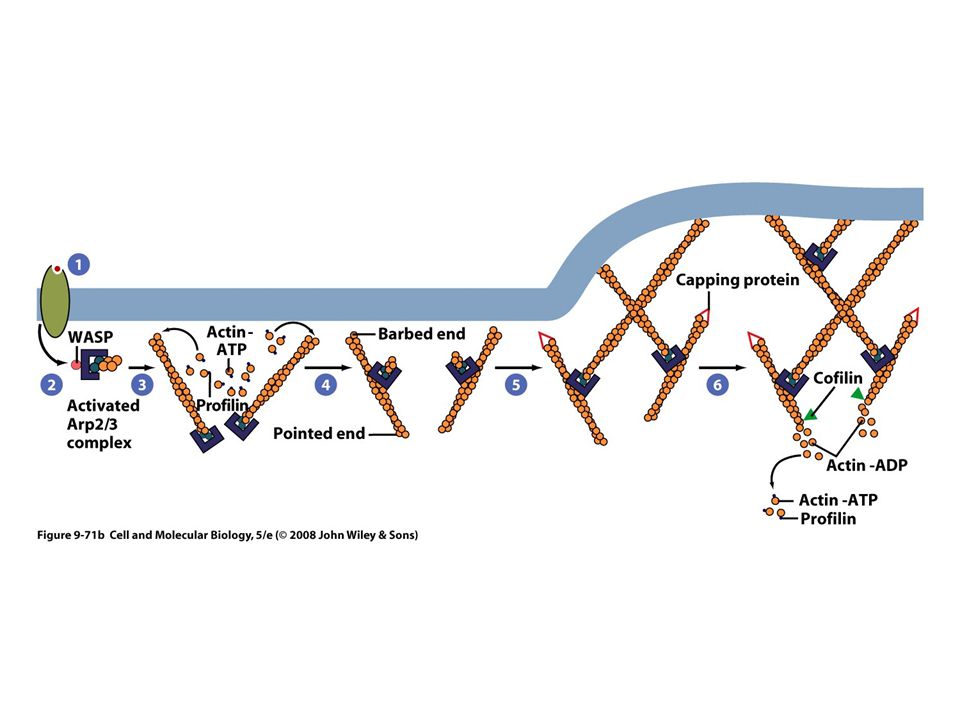
Proteínas de nucleación La nucleación requiere la unión de por lo menos dos o tres monómeros de actina en la orientación apropiada Las células tienen un complejo proteico el complejo Arp2/3 “Arp” actin-related proteins El complejo Arp2/3 debe activarse para comenzar la nucleación Formación de molde

69
Proteínas de unión a actina
Proteínas para secuestro de monómeros Timosinas, se unen con los monómeros de actinaG Proteínas para bloqueo de extremos (tapaderas) Regulan la longitud de los filamentos Proteínas polimerizadoras de monómeros Profilina Proteínas despolimerizadoras de actina Cofilina, locomoción celular y fagocitosis
 Regulan la longitud de los filamentos. Proteínas polimerizadoras de monómeros. Profilina. Proteínas despolimerizadoras de actina. Cofilina, locomoción celular y fagocitosis.")
70
Proteínas de unión a actina
Proteínas que forman enlaces cruzados Pueden alterar la organización tridimensional del filamento Proteínas cortadoras de filamentos Puede unirse con un filamento ya formado y romperlo en dos Proteínas de unión con membrana Vinculina, unión indirecta de los filamentos con la membrana plasmática

72
Timosina Arp2/3 Profilina Vinculina Filamina Fimbrina Cofilina

73
Locomoción celular Necesaria para muchas actividades en vertebrados superiores Desarrollo de órganos y tejidos Formación de vasos sanguíneos Desarrollo de axones Cicatrización de heridas Diseminación de tumores cancerosos

75
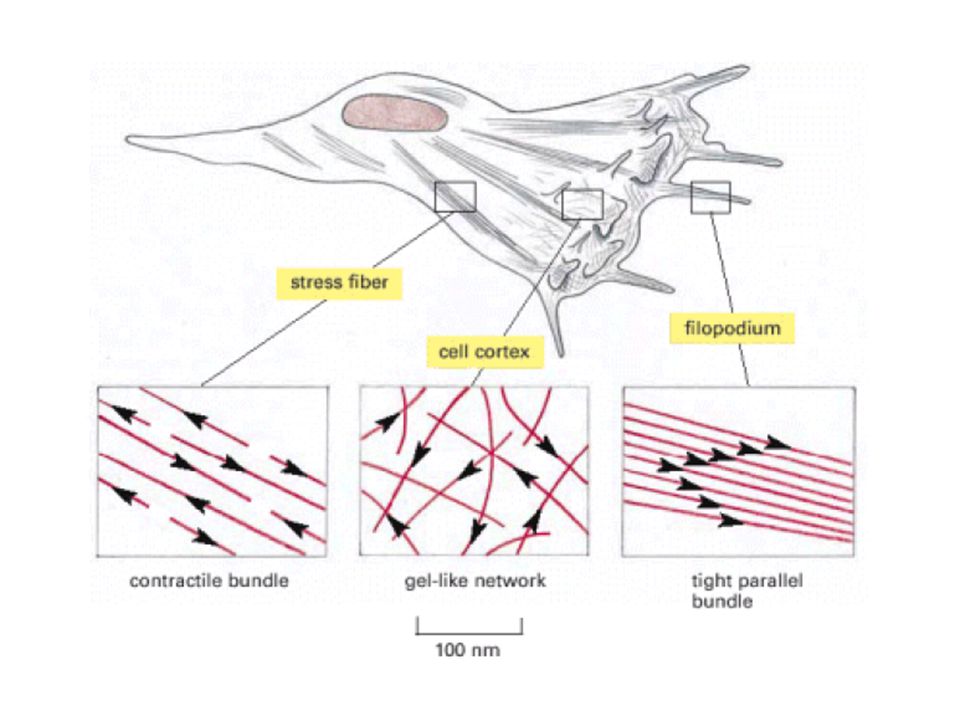
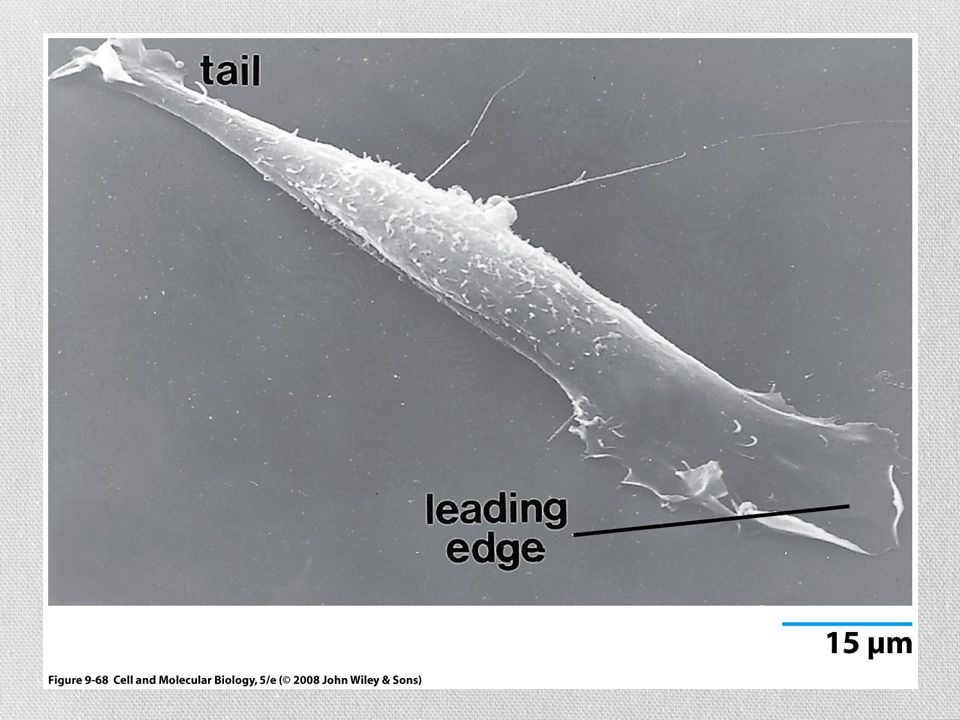
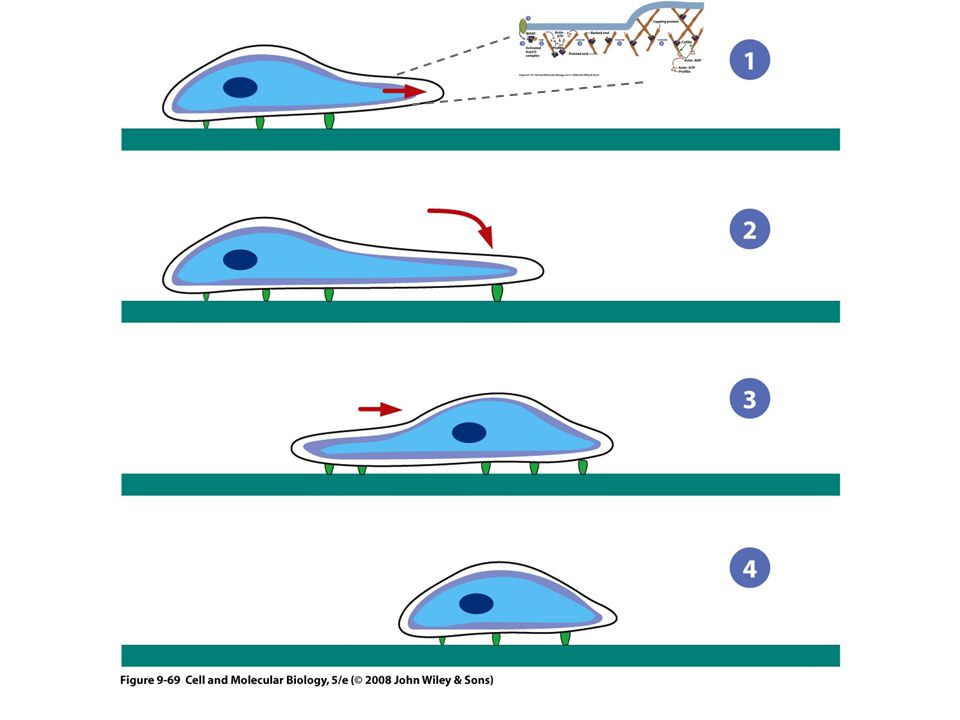
Locomoción celular El movimiento inicia por una protrusión de una parte de la superficie celular en la dirección en que la célula va a moverse Una parte de la superficie inferior de la protrusión se adhiere al sustrato y forma un anclaje temporal La mayor parte de la célula se impulsa hacia el frente sobre los contactos adhesivos, que al final se convierten en la parte posterior de la célula

76
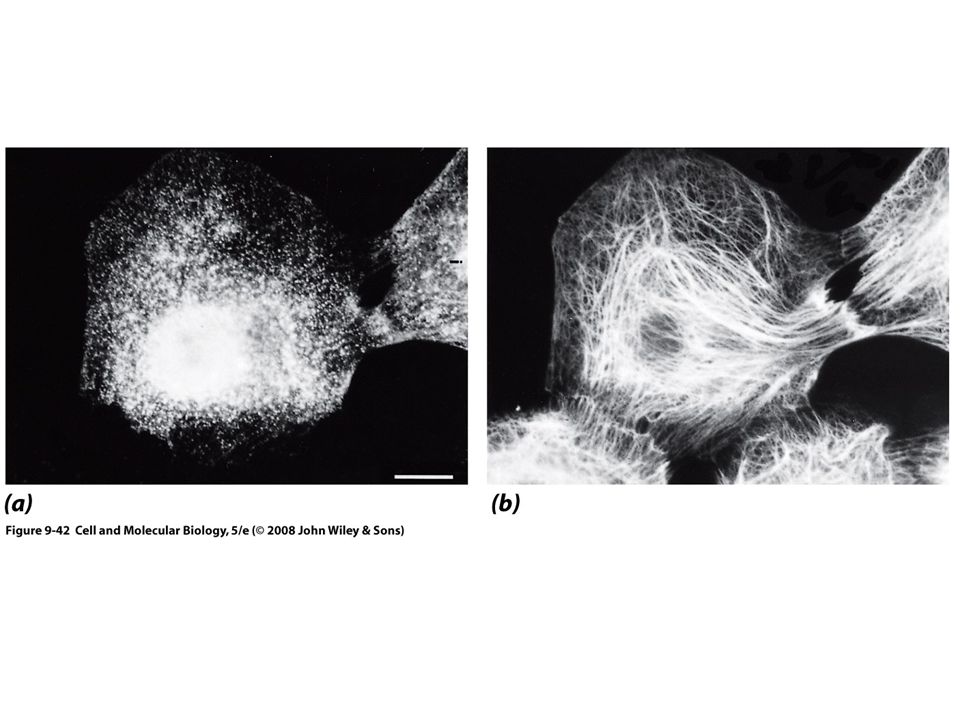
Locomoción celular 4) La célula rompe los contactos traseros con el sustrato, lo que causa la retracción del margen final o cola Lamelipodio Protrusión ancha, semejante a un velo

78
Locomoción celular Leucocito, recibe una señal química
Una vez recibido el estimulo, inicia la polimerización localizada de actina Tiene una familia de proteínas llamada WASP que activa el complejo Arp2/3 Síndrome de Wiskott-Aldrich

80
Lamelopodio

Presentaciones similares