Descargar la presentación
La descarga está en progreso. Por favor, espere

1
CITOESQUELETO

2
Citoplasma El citoplasma es el espacio celular comprendido entre la membrana plasmática y la envoltura nuclear. Está constituido por el citosol, el citoesqueleto y los orgánulos celulares.

3
Citosol El citosol (también llamado hialoplasma) es el medio interno del citoplasma. En él flotan el citoesqueleto y los ribosomas. Está formado por un 85% de agua con un gran contenido de sustancias dispersas en él de forma coloidal (prótidos, lípidos, glúcidos, ácidos nucleicos y nucleótidos así como sales disueltas. En él se produce una ingente cantidad de reacciones metabólicas importantes: glucólisis, gluconeogénesis, fermentación láctica, etc

4
CITOESQUELETO El citoesqueleto aparece en todas las células eucariotas. La composición química es una red de fibras de proteína (microfilamentos, filamentos intermedios y microtúbulos). Sus funciones son: Forma celular Soporte celular Movimiento celular Movimiento de organelos División celular
. Sus funciones son: Forma celular. Soporte celular. Movimiento celular. Movimiento de organelos. División celular.")
5
EL CITOESQUELETO FILAMENTOS PROTEICOS
se prolonga en consta de FILAMENTOS PROTEICOS CILIOS Y FLAGELOS MICROFILAMENTOS DE ACTINA FILAMENTOS INTERMEDIOS MICROTÚBULOS Intervienen en: morfología, estructura interna y movimiento

6
COMPOSICIÓN DEL CITOESQUELETO

10
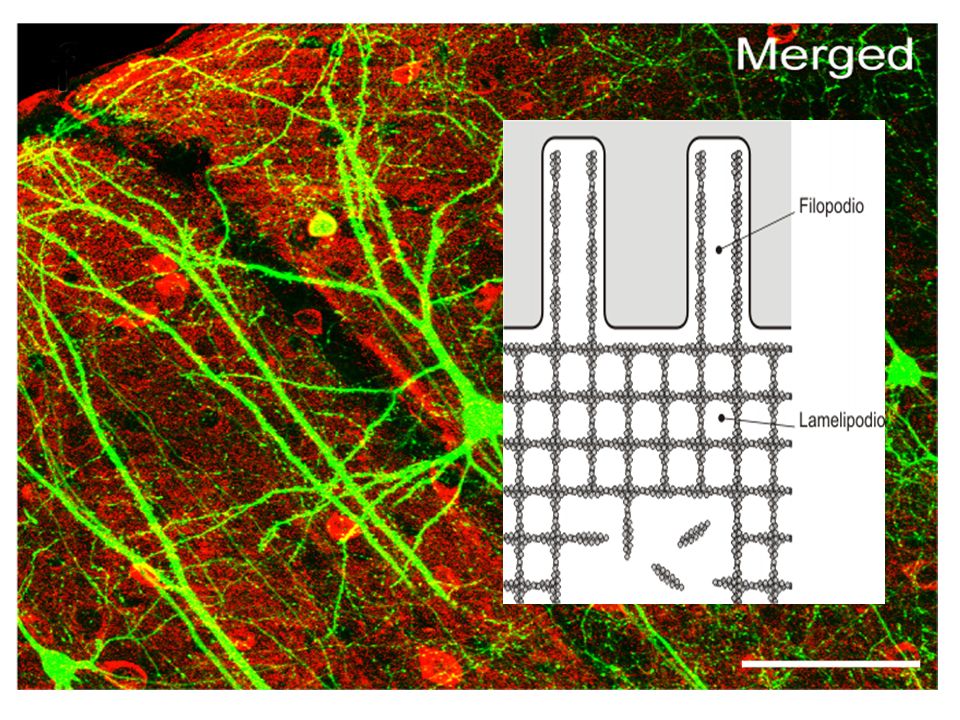
MICROFILAMENTOS Son las fibras más delgadas de 3-6 nm (nanómetros=milmillonésimas de metro= 10-9), están formados por la proteína actina. La actina es una proteína con funciones contráctiles, es también la proteína celular más abundante. La asociación de estos microfilamentos de actina con la proteína miosina es la responsable de la contracción muscular. Los microfilamentos también pueden llevar a cabo los movimientos celulares, incluyendo desplazamiento, contracción y citiocinesis.

11
ACTINA Los filamentos de actina poseen unos 7 nm de diámetro. Es el valor más pequeño dentro de los filamentos que componen el citoesqueleto. Poseen un extremo más y otro menos, es decir, son filamentos polarizados. El extremo más se denomina así porque en él predomina la polimerización, adición de nuevos monómeros, respecto a la despolimerización, mientras que en el extremo menos predomina la despolimerización. El mecanismo de crecimiento y acortamiento de la longitud de los filamentos de actina es por polimerización y despolimerización, respectivamente, de los monómeros que los componen. En la célula se crean y se destruyen filamentos de actina continuamente.

13
Los monómeros de actina se organizan gracias a las denominadas proteínas moduladoras de la actina, las cuales afectan a la velocidad de polimerización de los monómeros de actina, así como a la organización tridimensional de los filamentos. Estas proteínas moduladoras se pueden clasificar en diferentes tipos : La profilina, se unen a los monómeros de actina libres e impiden su unión a filamentos preexistentes, mientras otras, como la timosina, favorecen su unión. Hay proteínas moduladoras, como las fimbrina y la α-actinina, que permiten la formación de haces de filamentos de actina mediante el establecimiento de puentes cruzados entre filamentos, mientras otras, como la filamina, permiten la formación de estructuras reticulares. Ciertas proteínas moduladoras, como la cofilina, la katanina o la gesolina, provocan la rotura y remodelación de los filamentos de actina. También hay proteínas que median en la interacción de los filamentos de actina con otras proteínas relacionadas, como es el caso de la tropomiosina, que media la interacción entre actina y miosina

15
La actina es la proteína base de los microfilamentos.
El monómero es conocido como actina G, o actina globular. En presencia de ATP, se polimeriza formando largas hélices dobles, denominadas actina F, o actina filamentosa. Para que se lleve a cabo esta polimerización el ATP debe convertirse en ADP, liberando la energía necesaria para el proceso. La actina, presenta polaridad, tiende a polimerizarse (alargarse) y despolimerizarse (acortarse) a gran velocidad por un extremo más (el extremo positivo), y a realizar los mismos procesos por el otro extremo, menos (extremo negativo), a menor velocidad.
 y despolimerizarse (acortarse) a gran velocidad por un extremo más (el extremo positivo), y a realizar los mismos procesos por el otro extremo, menos (extremo negativo), a menor velocidad.")
16
Existen ciertos factores que condicionan la acción de estas proteínas moduladoras, como la variación en la concentración de calcio, proteínas como las Rho-GTPasas, la presencia de lípidos o la mayor o menor expresión génica de sus ARN mensajeros. También hay drogas que afectan a la polimerización de los filamentos de actina. Por ejemplo, las citocalasinas impiden la polimerización y las faloidinas impiden la despolimerización

17
FUNCIONES DE LA ACTINA Movimiento Endocitosis (fagocitosis) Citocinesis. El estrangulamiento final del citoplasma durante el proceso de división celular se produce gracias a un anillo de actina, que, ayudado por la miosinas, va estrechando su diámetro progresivamente hasta la separación completa de los dos citoplasmas de las células hijas. Contracción
 Citocinesis. El estrangulamiento final del citoplasma durante el proceso de división celular se produce gracias a un anillo de actina, que, ayudado por la miosinas, va estrechando su diámetro progresivamente hasta la separación completa de los dos citoplasmas de las células hijas. Contracción")
20
FILAMENTOS INTERMEDIOS
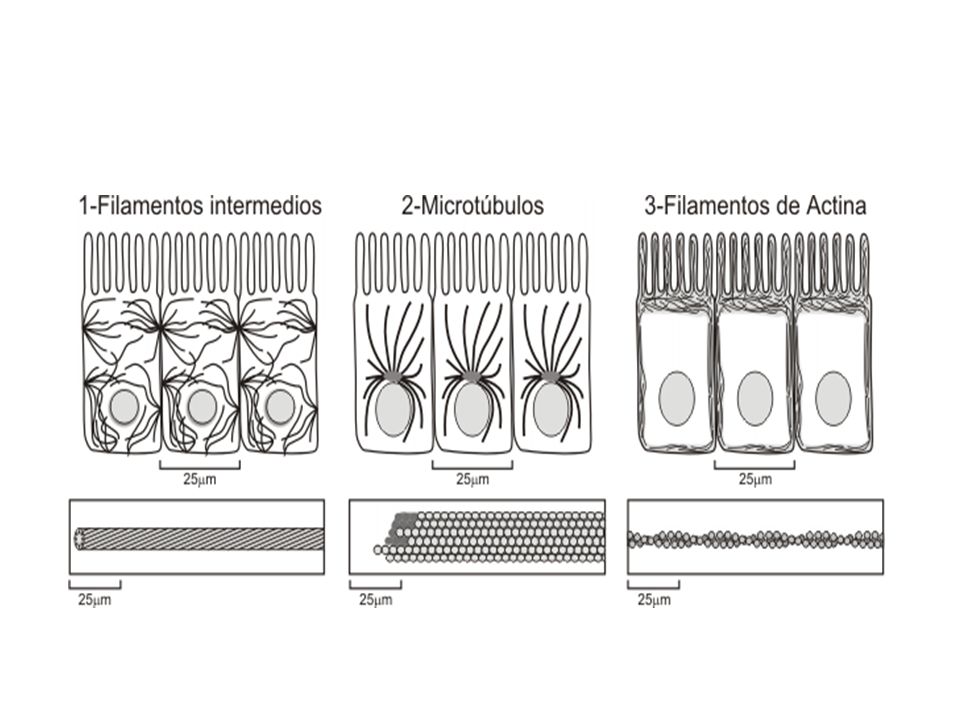
SON filamentos de diámetro mayor a la actina (10 nm) pero menor que los microtúbulos. CARACTERÍSTICAS Proteínas fibrosas, muy resistentes. Forman redes que parten del núcleo. En todas las células eucariotas.
 pero menor que los microtúbulos. CARACTERÍSTICAS. Proteínas fibrosas, muy resistentes. Forman redes que parten del. núcleo. En todas las células eucariotas.")
21
FILAMENTOS INTERMEDIOS
Los filamentos intermedios tienen 10 nm de diámetro y proveen fuerza de tensión (resistencia mecánica) a la célula. Según el tipo celular varían sus proteínas constitutivas. Podemos decir que existen seis tipos de filamentos intermedios Neurofilamentos (en la mayoría de las neuronas). Filamentos de desmina, en el músculo. Filamentos gliales, en las células del mismo nombre , que sirven de soporte en el cerebro, médula espinal y sistema nervioso periférico. Filamentos de vimentina en células del tejido conjuntivo y en los vasos sanguíneos. Queratinas epiteliales, (o filamentos de queratina o también llamados tonofilamentos), en células epiteliales. Laminofilamentos, forman la lámina nuclear, una delgada malla de filamentos intermedios sobre la superficie interna de la envoltura nuclear. Son los únicos que no se encuentran en el citoplasma.
 a la célula. Según el tipo celular varían sus proteínas constitutivas. Podemos decir que existen seis tipos de filamentos intermedios. Neurofilamentos (en la mayoría de las neuronas). Filamentos de desmina, en el músculo. Filamentos gliales, en las células del mismo nombre , que sirven de soporte en el cerebro, médula espinal y sistema nervioso periférico. Filamentos de vimentina en células del tejido conjuntivo y en los vasos sanguíneos. Queratinas epiteliales, (o filamentos de queratina o también llamados tonofilamentos), en células epiteliales. Laminofilamentos, forman la lámina nuclear, una delgada malla de filamentos intermedios sobre la superficie interna de la envoltura nuclear. Son los únicos que no se encuentran en el citoplasma.")
22
TIPO DE FILAMENTOS INTERMEDIOS
TIPOS FUNCIONES QUERATINA Mantenimiento de membranas (Tonofilamentos) Da estructura a la célula NEUROFILAMENTOS Prolongaciones neuronas OTROS Soportar tensiones Vimentina Desmina GFAP Células epiteliales Cel. Musculares lisas
 Da estructura a la célula. NEUROFILAMENTOS. Prolongaciones neuronas. OTROS. Soportar tensiones. Vimentina. Desmina. GFAP. Células epiteliales. Cel. Musculares lisas.")
23
Los filamentos intermedios son componentes del citoesqueleto, formados por agrupaciones de proteínas fibrosas. Su nombre deriva de su diámetro, de 10 nm, menor que el de los microtúbulos, de 24 nm, pero mayor que el de los microfilamentos, de 7 nm. A diferencia de los microfilamentos y microtúbulos, los filamentos intermedios al agruparse pierden polaridad, por lo tanto no presentan extremo + y extremo

24
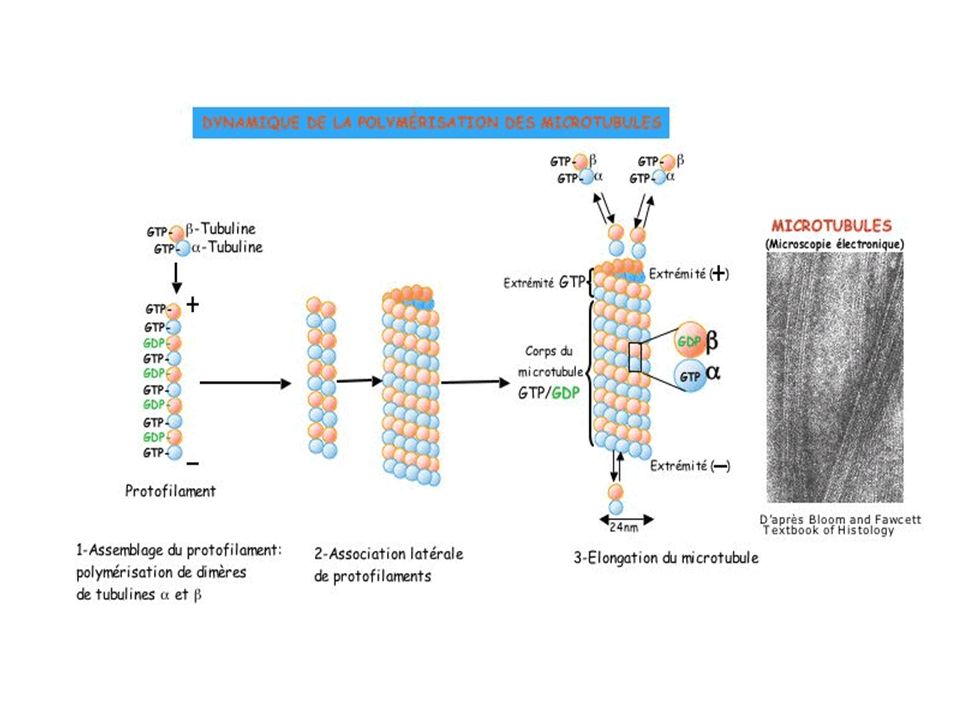
MICROTUBULOS Los microtúbulos son un elemento del citoesqueleto formado por dímeros de α- y β- tubulina que se organizan formando un tubo alargado. Son tubos largos y relativamente rígidos. Sus paredes están formados por unas subunidades proteicas globulares denominadas tubulinas. Éstas se asocian en dímeros compuestos de dos tipos de tubulinas: α y β Estas parejas se alinean ordenadamente, mediante enlaces no covalentes, en filas longitudinales que se denominan protofilamentos

25
Los microtúbulos son tubos cilíndricos de 20-25 nm de diámetro
Los microtúbulos son tubos cilíndricos de nm de diámetro. Están compuestos de subunidades de la proteína tubulina , estas subunidades se llaman alfa y beta.

26
PROTOFILAMENTOS Un microtúbulo tipo contiene trece protofilamentos.
Cada protofilamento tiene una polaridad estructural: la α-tubulina siempre formará un extremo del protofilamento y la β el otro. El microtúbulo es una estructura polarizada. Se denomina extremo más al extremo donde hay una α-tubulina y menos donde está la β-tubulina. Los nuevos dímeros de tubulina se añade con una mayor eficacia a la α-tubulina que a la β-tubulina, por lo que el extremo más es el lugar preferente de crecimiento y predomina la polimerización respecto a las despolimerización. En el extremo menos predomina la despolimerización respecto a la polimerización. Por ello los microtúbulos suelen crecer por el extremo más y, si no está protegido, decrecer por el extremo menos. Sin embargo, el extremo más es muy dinámico y en él se suceden procesos de polimerización y despolimerización, algunos tan drásticos que pueden hacer desaparecer por completo al microtúbulo.

27
INESTABILIDAD DINAMICA DEL MICROTUBULO
Una vez se ha producido el comienzo de la formación de un microtúbulo, la incorporación de nuevos dímeros de tubulina hace que el microtúbulo crezca en longitud. Este crecimiento a veces se detiene repentinamente y el microtúbulo comienza a despolimerizarse, llegando a veces incluso a desaparecer, o más frecuentemente reinicia el proceso de polimerización. A estas alternancias entre polimerización y despolimerización es a lo que se llama inestabilidad dinámica.

28
Los microtúbulos están continuamente polimerizando y despolimerizando, fundamentalmente en su extremo más

29
MTOCs Centros organizadores de microtúbulos. Estos son los lugares donde comienza la polimerización de un nuevo microtúbulo y donde suelen estar anclados sus extremos menos. El principal MTOC en las células animales es el centrosoma, el cual controla el número, localización y orientación de los microtúbulos en el citoplasma. Hay un centrosoma por célula, cuando ésta se encuentra en la fase G1 o G0 del ciclo celular, y se localiza cerca del núcleo. El centrosoma se compone de dos compartimentos: uno central formado por un par de centriolos dispuestos de forma ortogonal y otro periférico formado por material proteico denominado matriz pericentriolar. Los centriolos son estructuras cilíndricas formadas por 9 tripletes de microtúbulos formando sus paredes.

30
En la matriz pericentriolar hay numerosas moléculas entre las que se encuentra la γ- tubulina, las cuales forman unos anillos denominados anillos de γ-tubulina. Estos anillos actúan como molde y lugar de nucleación y anclaje de nuevos microtúbulos. Los centriolos, sin embargo, no desempeñan papel alguno en la polimerización y dirección de los microtúbulos. La misión de los centriolos es todavía un misterio puesto que las células vegetales carecen de ellos y no por eso dejan de dividirse u orientar sus microtúbulos. El centrosoma no sólo participa en la polimerización de los microtúbulos sino que también es importante en la regulación del ciclo celular por la presencia en la matriz pericentriolar de numerosas proteínas que afectan al avance del ciclo celular y por la organización del uso mitótico

33
FUNCION DE LOS MICROTUBULOS
Organización y movimiento de orgánulos Los microtúbulos se pueden clasificar en dos grandes grupos: Aquellos que son estables, presentes en los cilios y flagelos, Otros más dinámicos y cambiantes que se encuentran en el citoplasma Aparte del papel de los microtúbulos citoplasmáticos en el movimiento de los cromosomas, mediante la formación del huso mitótico, que se verá más adelante, participan en el movimiento de orgánulos como las mitocondrias, lisosomas, pigmentos, gotas de lípidos. Son también necesarios para dirigir el tráfico vesicular

34
Los microtúbulos son relativamente inertes en cuanto que no interaccionan directamente con los orgánulos. Los desplazamientos de orgánulos son producidos por una serie de proteínas especiales llamadas proteínas motoras. Estas proteínas pertenecen a dos familias: quinesinas y dineínas, las cuales se desplazan por el microtúbulo en direcciones opuestas: las quinesinas hacia el extremo más y las dineínas hacia el extremo menos. Tanto unas como otras tienen dos estructuras globulares y una cola. Las zonas globulares unen ATP e interaccionan con los microtúbulos con una orientación determinada, mientras que las colas se unen a las cargas que han de transportar. La cola es lo que determina qué elemento es el transportable. La hidrólisis del ATP en las zonas globulares provoca el cambio estructural de la proteína y su desplazamiento a lo largo del microtúbulo.

35
MICROTÚBULOS ESTRUCTURA En citoplasma, centriolos, cilios y flagelos.
13 protofilamentos a-tubulina y b-tubulina hueco interior

36
MICROTÚBULOS FUNCIONES Formación huso mitótico Transporte intracelular
Cilios Flagelos pseudópodos Movimiento celular

37
Cilios y flagelos Los movimientos celulares más frecuentes son: por cilios y flagelos (cortos o largos pelillos que vibran y agitan el medio), o por seudópodos (movimientos ameboideos) prolongando el citoplasma y modificando la forma de la membrana como si fueran dedos que se arrastran al resto de la célula. Existen estructuras celulares en las células animales, en los gametos de algunas especies vegetales y en organismos unicelulares que poseen haces de microtúbulos altamente organizados y muy estables en cuanto a su disposición y longitud: los centriolos, los cilios y los flagelos. En esta sección vamos a estudiar a los cilios y a los flagelos.
, o por seudópodos (movimientos ameboideos) prolongando el citoplasma y modificando la forma de la membrana como si fueran dedos que se arrastran al resto de la célula. Existen estructuras celulares en las células animales, en los gametos de algunas especies vegetales y en organismos unicelulares que poseen haces de microtúbulos altamente organizados y muy estables en cuanto a su disposición y longitud: los centriolos, los cilios y los flagelos. En esta sección vamos a estudiar a los cilios y a los flagelos.")
38
APARATO CILIAR Los cilios son expansiones celulares filiformes, de unos 0,25 µm de diámetro y unos 10 a 15 µm de longitud, que aparecen en las células animales y en algunos protozoos. Suelen disponerse densamente empaquetados, a modo de cesped, en las superficies libres de numerosas células, como las que forman los epitelios de los tractos respiratorios, de los conductos del aparato reproductor femenino de mamíferos o de las branquias de los peces. Aparecen en numerosos protozoos. Son estructuras que pueden moverse y su principal misión es la de desplazar fluidos.

Presentaciones similares