Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Villanueva Alonso, Óscar Calvo Morales, Maria Dolores
SUPERCOMPLEJOS Villanueva Alonso, Óscar Calvo Morales, Maria Dolores

2
La mitocondria

3
DNA mitocondrial El mtDNA es una molécula de doble hebra de DNA circular El mtDNA no tiene intrones y los tRNAs sirven como elementos de procesamiento Tiene sus propios ribosomas El mtDNA posee su propio código genético

4
Características del mtDNA
Herencia materna Las mitocondrias se heredan exclusivamente de la madre Poliplasmia Cada célula con entre 100 y copias de mtDNA dependiendo del tejido Alta tasa de mutación El mtDNA presenta una tasa de mutación espontánea 10 veces superior a la del DNA nuclear Segregación mitótica Homoplasmia Heteroplasmia

5
Dificultades de la genética mitocondrial en animales
El mtDNA no es manipulable “in vivo”: No se pueden transformar mitocondrias No se puede realizar mutagénesis dirigida. El mtDNA de una especie es incompatible con el nDNA de otra. No se pueden hacer estudios de genética funcional Dificultad en la generación de modelos animales de patologías mitocondriales

6
RESPIRACIÓN AEROBIA: cadena de transporte de electrones

7
Solo unas pocas subunidades de los complejos respiratorios están codificados por el mtDNA

8
Estructura de los complejos

9
MODELOS: “estado líquido” “estado sólido”
todos los componentes de la cadena de electrones difunden libremente en la membrana interna de la mitocondria y la transferencia de electrones entre unos y otros depende de encuentros a azar “estado sólido” los complejos están asociados permanentemente formando supercomplejos

10
Ventajas de los supercomplejos
Estabilización de la estructura de los compuestos individuales Canalización de los reactivos intermedios La ubiquinona y el citocromo c pasan directamente de un complejo al siguiente Secuestro de las especies intermedias Se evita que puedan reaccionar con el O2 para dar radicales libres

11
b) supercomplejo I1III2IV1
a) supercomplejo I1III2 b) supercomplejo I1III2IV1 Schägger et Peiffer (2001) JBC, 276,
 supercomplejo I1III2. b) supercomplejo I1III2IV1. Schägger et Peiffer (2001) JBC, 276,")
12
Modelos para la estequiometria de los complejos
Schägger et Peiffer (2001) JBC, 276,
 JBC, 276,")
13
Estudio de la interrelación entre el CI y el CIII

14
Observaciones Pacientes con intolerancia al ejercicio con deficiencias en el CIII en el tejido muscular Deficiencias en el CIII pueden que pueden cursar solas pero suelen hacerlo como deficiencias combinadas en el CIII+CI No existe simetria el fenotipo de las deficiencias en el CI que suelen cursar aisladas La mayoría de las deficiencias en el CIII se deben a mutaciones en el gen mitocondrial del CIII, el gen del cit b

15
Línea celular de ratón L929 Obtención de la linea A22
Glu 373 Lys

16
Creación de la línea transmitocondrial FA22
A22 y FA22 son homoplásmicas para la mutación en el cit b No se observa actividad enzimática del CIII ni del CI

17
A22 tiene el mismo fenotipo que las deficiencias combinadas del CIII + CI debidas a mutaciones en el cit b

18
El complejo III no se forma en A22 ni en FA22
Los niveles de complejo I en A22 y FA22 están seriamente reducidas

19
Existe interdependencia en la formación/estabilidad entre los 2 complejos

20
Inhibimos el CIII de forma farmacológica
Después de 2 semanas de tratamiento de de las células con el inhibidor específico de la actividad del CIII, el CI no esta afectado

21
Es la ausencia física del CIII y no la pérdida de su actividad lo que produce la pérdida del CI

22
¿Es necesario el CIII para la formación del CI o solo lo estabiliza?
En las células control el CI aumenta con el tiempo, en cambio en las A22 el CI disminuye a medida que pasa el tiempo

23
En ausencia del CIII el CI si que se forma, pero es inestable

24
Estudio de la biopsia muscular de un paciente con mutación en el cit b
Se produce la pérdida concomitante del CIII y el CI

25
La presencia del CIII es requerida para la estabilidad del CI en células humanas diferenciadas

26
Explicación del fenotipo de pacientes con mutaciones en el CIII
CIV CIII CV Pacientes tipo A Pacientes tipo B El fenotipo de los pacientes estará modulado por el grado de heteroplasmia de la mutación

27
La existencia de la asociación entre los dos complejos respiratorios, ya documentada, es coherente con estos descubrimientos

28
ESTUDIO DE LA ARQUITECTURA DE SUPERCOMPLEJOS MEDIANTE MICROSCOPÍA ELECTRÓNICA

29
Para la caracterización bioquímica y estructural
(a), BN-PAGE (poliacrilamida gradiente de concentración T: El 3-13 %) de mitocondrias de corazón bovinas solubilizadas. (b), BN-PAGE (T: El 3-5 %). Ambas tiras de supercomplejos estaban en la proximidad cercana en una BN-PAGE (la T: El 3-13 %).
, BN-PAGE (poliacrilamida gradiente de concentración T: El 3-13 %) de mitocondrias de corazón bovinas solubilizadas. (b), BN-PAGE (T: El 3-5 %). Ambas tiras de supercomplejos estaban en la proximidad cercana en una BN-PAGE (la T: El 3-13 %).")
30
La separación fue mejorada en una BN-PAGE con la T: El 3-5 %, BN- PAGE (T: El 3-13 %) de los supercomplejos aislados I1III2 y I1III2IV1. El Supercomplejo I1III2IV1 (BANDA 2) está intacto después de la electroelución, pero el supercomplejo I1III2 (BANDA 1) muestra dos bandas adicionales muy débiles de I y III2.
 está intacto después de la electroelución, pero el supercomplejo I1III2 (BANDA 1) muestra dos bandas adicionales muy débiles de I y III2.")
31
Para la actividad enzimática
La actividad del complejo NADH deshidrogenasa fue mostrada por bandas púrpuras que son resultado de la precipitación en gel de formazan

32
El la actividad de la citocromo c oxidasa del complejo IV fue determinada por la oxidación del citocromo c dependiente de 3,3 '-diaminobenzidina al óxido marrón e indamina precipitada

33
La actividad del complejo I en el supercomplejo I1III2 y I1III2IV1 así como la actividad del complejo IV para el supercomplejo I1III2IV1 en las muestras aisladas fue demostrada:

34
Ensayos espectofotométricos para evaluar la actividad de la NADH:UBIQUINOL reductasa del complejo I y la actividad de la citocromo c reductasa del complejo III en las muestras electroeluidas. En las condiciones usadas, se obtuvo: Para concluir ambos supercomplejos aislados mostraron actividad, pero el supercomplejo I1III2IV1 era considerablemente más activo.

35
Para la microscopía de electrones
I1III2 I1III2IV1 Las imágenes triangulares están indicadas por círculos rojos. Las formas L por círculos amarillos. Los círculos verdes en (a) indican las partículas que podrían ser el complejo dimérico III proveniente de la disociación del supercomplejo I1III2. Las barras de escala representan 100 nm.
 indican las partículas que podrían ser el complejo dimérico III proveniente de la disociación del supercomplejo I1III2. Las barras de escala representan 100 nm.")
36
Las dos vistas representan las dos orientaciones preferenciales de los supercomplejos.
Una orientación es preferida sobre la otra.

37
Los mapas de proyección hechos en promedio de los supercomplejos.
(a) vistas superiores (espacio intermembrana) del supercomplejo I1III2IV1 (con un promedio de 228 partículas). (b) el Supercomplejo I1III2 encabezan la vista el promedio de 66 partículas). (c) mapa diferencial entre I1III2IV1 y I1III2. (d) vista superior del espacio de la matriz de I1III2IV1. (e) Vista lateral de supercomplejo I1III2IV1 (promedio de 70 partículas). (f) la vista superior del complejo bovino IV a través de rayos X, visto desde el espacio intermembrana.
 vistas superiores (espacio intermembrana) del supercomplejo I1III2IV1 (con un promedio de 228 partículas). (b) el Supercomplejo I1III2 encabezan la vista el promedio de 66 partículas). (c) mapa diferencial entre I1III2IV1 y I1III2. (d) vista superior del espacio de la matriz de I1III2IV1. (e) Vista lateral de supercomplejo I1III2IV1 (promedio de 70 partículas). (f) la vista superior del complejo bovino IV a través de rayos X, visto desde el espacio intermembrana.")
38
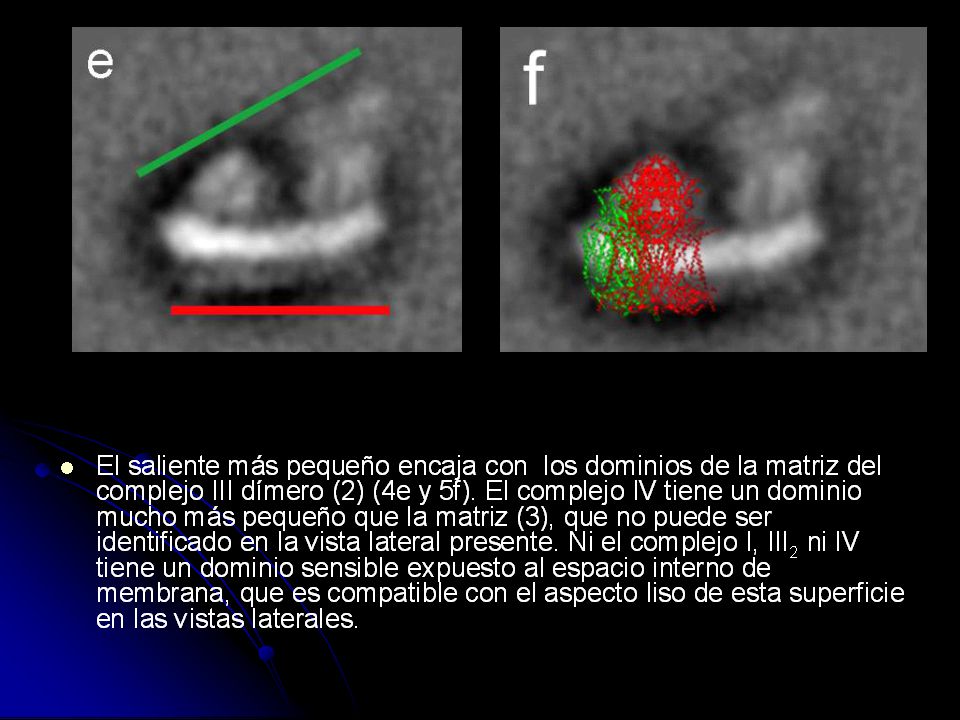
300 partículas observada con menos frecuencia que las anteriores fueron seleccionadas. Después de la alineación y haciendo un promedio, la imagen mostró dos salientes grandes sobre un lado de la línea brillante, mientras que otra superficie apareció lisa.

39
Si comparamos con un mapa de proyección de tinción negativa del complejo I en corazón bovino (a) se identifica el saliente más grande (e) como el brazo de la matriz de complejo I. El dominio de la membrana del complejo I en el supercomplejo se extiende a lo largo del saliente más pequeño en esta vista. La línea brillante alrededor del supercomplejo puede ser interpretada como los dominios transmembrana de la proteína debido al detergente y posiblemente la mancha sea debida al azul de Coomassie de la BN-PAGE.

41
La asimetría de la vista lateral (e) claramente explica las distintas vistas triangulares (a y d), que puede ser interpretado como la vista perpendicular a la membrana del lado de la matriz y el espacio intermembrana, respectivamente.
 claramente explica las distintas vistas triangulares (a y d), que puede ser interpretado como la vista perpendicular a la membrana del lado de la matriz y el espacio intermembrana, respectivamente.")
42
(a) proyección de la estructura del complejo I.
Las disposiciones alternativas de los complejos I y III2 en supercomplejo I1III2IV1. (a) proyección de la estructura del complejo I. (b) las vistas laterales de las estructuras de rayos X del complejo IV (verde) y (c) el complejo III dimérico (rojo) dibujado en la misma escala que los mapas de proyección. (d) Estructura de rayo X de complejo IV complejo (verde) (e) complejo III2 (rojo), ambos vistos desde el espacio intermembrana.
 proyección de la estructura del complejo I. (b) las vistas laterales de las estructuras de rayos X del complejo IV (verde) y (c) el complejo III dimérico (rojo) dibujado en la misma escala que los mapas de proyección. (d) Estructura de rayo X de complejo IV complejo (verde) (e) complejo III2 (rojo), ambos vistos desde el espacio intermembrana.")
43
(f) las estructuras de rayo X de complejo III2 y IV superpuesto sobre el mapa de proyección del supercomplejo I1III2IV1 (vista lateral).
 las estructuras de rayo X de complejo III2 y IV superpuesto sobre el mapa de proyección del supercomplejo I1III2IV1 (vista lateral).")
44
[contacto con el complejo I y IV, menos contacto con III2]
(g) Modelo 1: El complejo III2 y IV superpuesto sobre el supercomplejo I1III2IV1 (como mirando desde el espacio intermembrana). [contacto con el complejo I y IV, menos contacto con III2]
![[contacto con el complejo I y IV, menos contacto con III2]](http://slideplayer.es/slide/5332130/17/images/44/%5Bcontacto+con+el+complejo+I+y+IV%2C+menos+contacto+con+III2%5D.jpg "(g) Modelo 1: El complejo III2 y IV superpuesto sobre el supercomplejo I1III2IV1 (como mirando desde el espacio intermembrana). [contacto con el complejo I y IV, menos contacto con III2]")
45
(h) Modelo 2: El complejo III2 está en la parte superior del supercomplejo. La masa restante en la parte inferior es el brazo de la membrana de complejo I. Así el complejo IV comparte una superficie de contacto grande con III2, pero hay poco contacto con el complejo I.

46
Vista superior del supercomplejo:
Modelo para la estructura del supercomplejo I+III2 de Arabidopsis thaliana, incorporando la estructura de rayos X del complejo III y la densidad de 3D de EM del complejo I del corazón bovino. Vista superior del supercomplejo: complejo I (amarillo). la otra mitad integrada por la membrana inferior de complejo III (la estructura que llena espacio) visto desde el lado de la matriz. Densidad de las proyecciones adicionales del complejo I se representan en color naranja. El contorno de los dominios hidrófilos de complejo III dentro de la matriz son indicadas por una línea de puntos verde.
. la otra mitad integrada por la membrana inferior de complejo III (la estructura que llena espacio) visto desde el lado de la matriz. Densidad de las proyecciones adicionales del complejo I se representan en color naranja. El contorno de los dominios hidrófilos de complejo III dentro de la matriz son indicadas por una línea de puntos verde.")
47
Complejo I (amarillo) y complejo III (verde), visto a un ángulo de 45 ° desde el plano de la membrana. Los componentes de los complejos I y III y de la membrana fuera del supercomplejo son indicados en azul.

48
CONTINUARÁ …

Presentaciones similares