Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Axón de jibia CURSOS_2010 FISIOLOGÍA GENERAL Hoy les hablara Carlos 15 de abril de 2010

2
Eliminando la variable espacio
Vm Electrodo que mide potencial Alambre axial La placa de tierra asegura que la resistencia del medio externo sea muy chica. Ro = 0 ¿Qué valor toma la constante de espacio? El alambre axial asegura que la resistencia del medio interno sea muy chica. Ri = 0.

3
Space clamp Generador de corriente Vm
A todo lo largo del axón pasa exactamente lo mismo, esto simplifica el análisis. Desaparece la variable espacio (Space clamp). El potencial de acción no se propaga. Se llama potencial de acción de membrana.
. El potencial de acción no se propaga. Se llama potencial de acción de membrana.")
4
Space clamp Generador de corriente Vm
Pero si queremos medir en forma correcta el potencial de membrana tenemos un problema porque mientras esté circulado corriente por el electrodo de tierra no sabemos el potencial eléctrico en la superficie externa de la membrana. Necesitamos otro electrodo para medir el potencial extracelular.

5
Potencial de acción de membrana

6
Potencial de acción de membrana

7
IIones, mAcm-2 I R V = 0 V = 0-IR Electrodo de tierra Superficie de la membrana Potencial eléctrico intracelular Vi, mV Intensidad de las corrientes iónicas IIones, mAcm-2 t, ms

8
Potencial eléctrico intracelular
Vi, mV t, ms RoIIones, Vo, mV Potencial eléctrico extracelular t, ms El potencial de acción aparece en el medio extracelular cono una espiga negativa y de pequeña amplitud. Esta es la base de los registros extracelulares de la actividad neuronal.

9
Space clamp Generador de corriente Vm Ve

10
Medida de Vm Vi, mV t, ms Vi Ve, mV Ve t, ms

11
Medida de Vm Vi -Vi Ve 0 V R R
Ver en einstein.ciencias.chile.cl Fisiologia General 2008, Lecturas, Circuitos con amplificadores operacionales.

12
Medida de Vm 0 V ? R R R R Vm Vi (t) R Vi -Ve -Vi Ve (t)
 R Vi -Ve -Vi Ve (t)")
13
Current clamp Generador de corriente Vm
Con este circuito puedo controlar la corriente y medir el potencial. Current clamp.

14
Voltage clamp Vm Vcomando
El potencial de la membrana es igual al potencial de comando en todos los puntos a lo largo del axón. El amplificador se encarga de pasar tanta corriente como sea necesario para mantener esta igualdad. Con este circuito puedo controlar el potencial. ¿Cómo podría medir la corriente?

15
Voltage clamp Vm Rf Vcomando -IRf
Con este circuito puedo controlar el potencial y medir la intensidad de la corriente. Voltage clamp. El potencial de la membrana es igual al potencial de comando en todos los puntos a lo largo del axón. El amplificador se encarga de pasar tanta corriente como sea necesario para mantener esta igualdad.

16
Voltage clamp Vm Rf Vcomando -IRf
Con este circuito puedo controlar el potencial y medir la intensidad de la corriente. Voltage clamp. El potencial de la membrana es igual al potencial de comando en todos los puntos a lo largo del axón. El amplificador se encarga de pasar tanta corriente como sea necesario para mantener esta igualdad.

17
? Rm Cm Vr Voltage clamp de un circuito pasivo Intracelular Vm Ii IC
Im Im ? Extracelular Tiempo

18
Rm Cm Vr Voltage clamp de un circuito pasivo Intracelular Ii IC Vm Vr
Im Extracelular Im Tiempo

19
X X Rm Cm Vr Eliminando la corriente capacitiva Intracelular Ii IC Vm
Extracelular Tiempo Im

20
Voltage clamp, resta de la corriente capacitiva
Vm Rf 0 V V0 = -IiRf Vcomando R R Cm Ajustar hasta que se cancele la corriente capacitiva -Vcomando

21
Voltage clamp de un axón de jibia
I, mA/cm2 t, ms -60,8mV 0mV

22
0mV -60,8mV TTX I, mA/cm2 t, ms Control
Voltage clamp de un axón de jibia en presencia de TTX, bloqueador de los canales de Na t, ms -60,8mV 0mV TTX I, mA/cm2 Control

23
La corriente de Na es la diferencia de la corriente control – corriente con TTX.
-60,8mV 0mV t, ms I, mA/cm2

24
Canales de potasio del axón de jibia

25
El curso temporal del desarrollo de la corriente de potasio no sigue un curso exponencial.
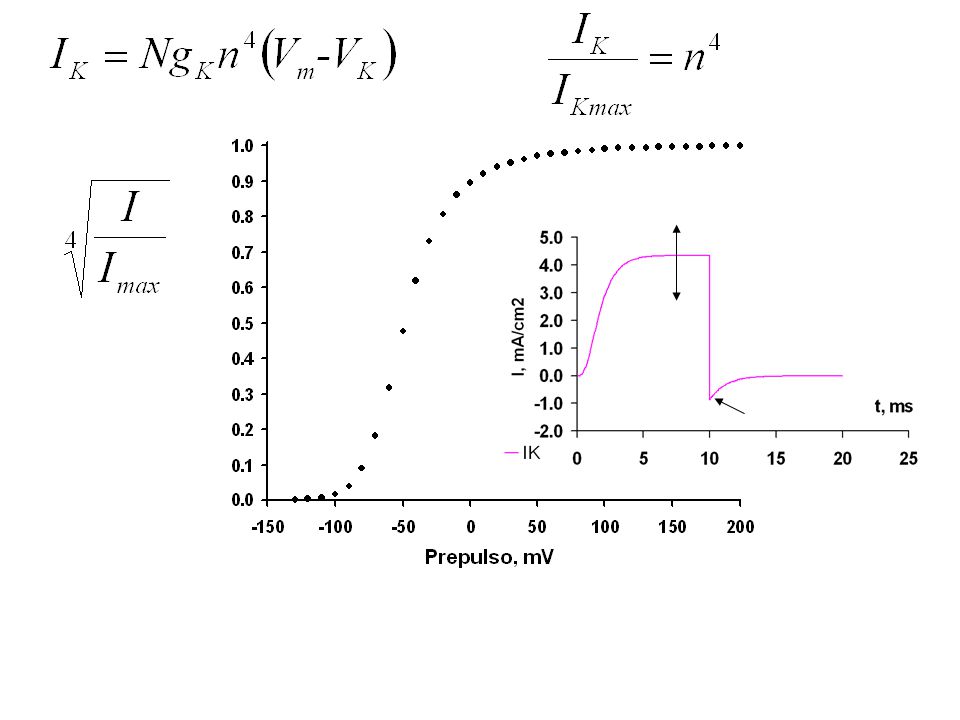
Pulso -120 a 100 mV Para estudiar la relación entre la intensidad de la corriente de potasio y el potencial de la membrana se mide a diferentes potenciales. Vm

26
IK, mA cm-2 Vm, mV Conductancia GK? Potencial de inversión, VK?
Relación entre la intensidad de la corriente de potasio y el potencial de la membrana. Conductancia GK? IK, mA cm-2 Potencial de inversión, VK? Vm, mV

27
? IK, mA cm-2 Vm, mV Conductancia GK? Potencial de inversión, VK?
Relación entre la intensidad de la corriente de potasio y el potencial de la membrana. Conductancia GK? Potencial de inversión, VK? IK, mA cm-2 VK = mV GK = 43.4 mS cm-2 Vm, mV ? -39.1 mV

28
? IK, mA cm-2 Vm, mV Conductancia GK? Potencial de inversión, VK?
Relación entre la intensidad de la corriente de potasio y el potencial de la membrana. Conductancia GK? Potencial de inversión, VK? IK, mA cm-2 VK = mV GK = 43.4 mS cm-2 Vm, mV ? -39.1 mV

29
IK, mA cm-2 Vm, mV Conductancia GK? Potencial de inversión, VK?
Relación entre la intensidad de la corriente de potasio y el potencial de la membrana. Conductancia GK? Potencial de inversión, VK? IK, mA cm-2 Vm, mV

30
Estrategia para buscar el potencial de inversión VK.
Vm, mV IK, mA/cm2 Estrategia para buscar el potencial de inversión VK. Sospecha: VK = -94 mV Problema: alrededor de -94 mV no se registra inversión de la corriente. ¿Por qué? Porque los canales están cerrados a -94 mV. Solución: 1) Abrir los canales, despolarizando la membrana 2) Cambiar el potencial en forma instantánea y 3) Medir la corriente antes de que los canales se cierren 4) Explorar la superficie I/V.
 Abrir los canales, despolarizando la membrana 2) Cambiar el potencial en forma instantánea y 3) Medir la corriente antes de que los canales se cierren 4) Explorar la superficie I/V.")
31
1) Abrir los canales, despolarizando la membrana 2) Cambiar el potencial en forma instantánea y 3) Medir la corriente antes de que los canales se cierren 4) Explorar la superficie I/V. Prepulso constante Pulso de prueba variable Vm

32
VK = -73,4 mV

33
Tarea: Calcular GK/GNa para estos canales
Sorpresa: El valor encontrado, -74 mV es menos negativo que el esperado según Nernst (-94mV). Una explicación posible: los canales de K no son idealmente selectivos: pasa algo de Na. VK = -73,4 mV Tarea: Calcular GK/GNa para estos canales El potencial de reposo es -60 mV, los potenciales de inversión de las corrientes de potasio y leak son -73 y -50 mV respectivamente. Calcule la razón GK/GLeak para el potencial de reposo.
. Una explicación posible: los canales de K no son idealmente selectivos: pasa algo de Na. VK = -73,4 mV. Tarea: Calcular GK/GNa para estos canales. El potencial de reposo es -60 mV, los potenciales de inversión de las corrientes de potasio y leak son -73 y -50 mV respectivamente. Calcule la razón GK/GLeak para el potencial de reposo.")
34
IK, mA/cm2 Vm, mV VK=-73.4 mV

35
IK, mA/cm2 Vm, mV VK=-73.4 mV

36
SEMINARIOS DE ESTUDIANTES
AVISO: SEMINARIOS DE ESTUDIANTES Forme un grupo de tres estudiantes para la presentación de su seminario. Cada grupo debe mandar una carta a declarando el artículo escogido, los integrantes del grupo, y la dirección electrónica de contacto del grupo. Antes de elegir el paper, revise en la página del curso ( ) el archivo LISTA DE PAPERS PARA SEMINARIOS, en que están indicados los papers ya seleccionados y aquellos aún disponibles. En la página del curso ( ) hay una carpeta que contiene las versiones PDF de todos los papers para seminarios. Si Ud. encontrara un paper que le interese y que no esté incluido en la lista, puede proponérselo al coordinador del curso y será incorporado si se considera adecuado. Espere confirmación a vuelta de correo. 19 de Abril de 2010
 el archivo LISTA DE PAPERS PARA SEMINARIOS, en que están indicados los papers ya seleccionados y aquellos aún disponibles. En la página del curso ( ) hay una carpeta que contiene las versiones PDF de todos los papers para seminarios. Si Ud. encontrara un paper que le interese y que no esté incluido en la lista, puede proponérselo al coordinador del curso y será incorporado si se considera adecuado. Espere confirmación a vuelta de correo. 19 de Abril de")
37
Continuación de la clase de canales de potasio del axón de jibia.
19 de Abril de 2010

38
Determinación de la conductancia del potasio a partir de la intensidad instantánea de la corriente de cola. Prepulso a 200 mV Pulse de prueba -100mV La conductancia se puede medir de la intensidad de la corriente instantánea de la cola, dividida por el potencial del pulso de prueba menos el potencial de inversión de la corriente.

39
Intensidad instantánea de la corriente de cola medida a -100 mV después de prepulsos de diferentes voltajes.

40
pK(v,) es la probabilidad de encontrar abierto el canal de potasio, al potencial del prepulso, y en estado estacionario. NB. Para sacar pK(v,) del análisis de las corrinentes de cola, no hace falta conocer el potencial de inversión de la corriente.
 del análisis de las corrinentes de cola, no hace falta conocer el potencial de inversión de la corriente.")
41
Los canales de K son tetrámeros.
Cada monómero tiene su propio sensor de potencial: es el segmento S4 que tiene varios residuos de arginina. Los sensores de potencial tiene dos estados: reposo y activo Los sensores de potencial operan en forma independiente. El canal se abre sólo cuando los cuatro sensores de potencial están activos. Sea n la probabilidad de encontrar un sensor activo La probabilidad de encontrar un canal abierto es... n4

42
NB. n es función del voltaje y del tiempo
NB. n es función del voltaje y del tiempo. n sólo es función del voltaje. Es el valor de n en estado estacionario: cuando se aplica un potencial por mucho tiempo.

44
V0 = -46 mV z = 1.4

45
V0 = -47 mV z = 1.8 Errores pesados por 1/n

46
V0 = -48 mV z = 1.2 Errores pesados por n La curva n vs Vm no se ajusta bien a una función de Boltzmann.

47
Cinética Caso n(0)=0, el curso temporal de la corriente es sigmoideo
Caso n(∞)=0, el curso temporal de la corriente es exponencial
=0, el curso temporal de la corriente es exponencial.")
48
Determinación de n. Vm 0 mV -100mV -140mV

49
Paso desde -140 mV a 0 mV Paso desde 0 mV a -100 mV

50
= 1.8 ms Paso desde -140 mV a 0 mV Paso desde 0 mV a -100 mV

51
= 1.8 ms = 4.7 ms Paso desde -140 mV a 0 mV Paso desde 0 mV a -100 mV

52
La constante de relajación n en función del potencial

53
Para el caso x = 0.5

56
La curva n vs Vm no se ajusta bien a una función secante hiperbólica.

57
Cálculo de a y b a partir de Po() y t
Las constantes alfa y beta se pueden calcular conociendo la probabilidad del canal abierto en estado estacionario y la constante de relajación

58
n n ms-1 Calcule n para el límite Vm-50 mV
Ecuaciones empíricas de Hodkin y Huxley Ver “El impulso nervioso” por F. Bezanilla en libro Biofísica y Fisiología Celular. Eds. R.Latorre et al.

59
n n Descripción completa de los canales de K del axón de jibia
Ngk = 35.3 mS/cm2 VK = -73,4 mV

60
Calcular la fracción de canales de potasio abiertos si se mantiene la membrana por mucho tiempo a -60 mV. Calcular la fracción de canales de potasio abiertos después de 5 ms de despolarizar desde -60 mV a 0 mV. Calcular la fracción de canales de potasio abiertos después de 1 ms de repolarizar desde 0 mV a -60 mV.

61
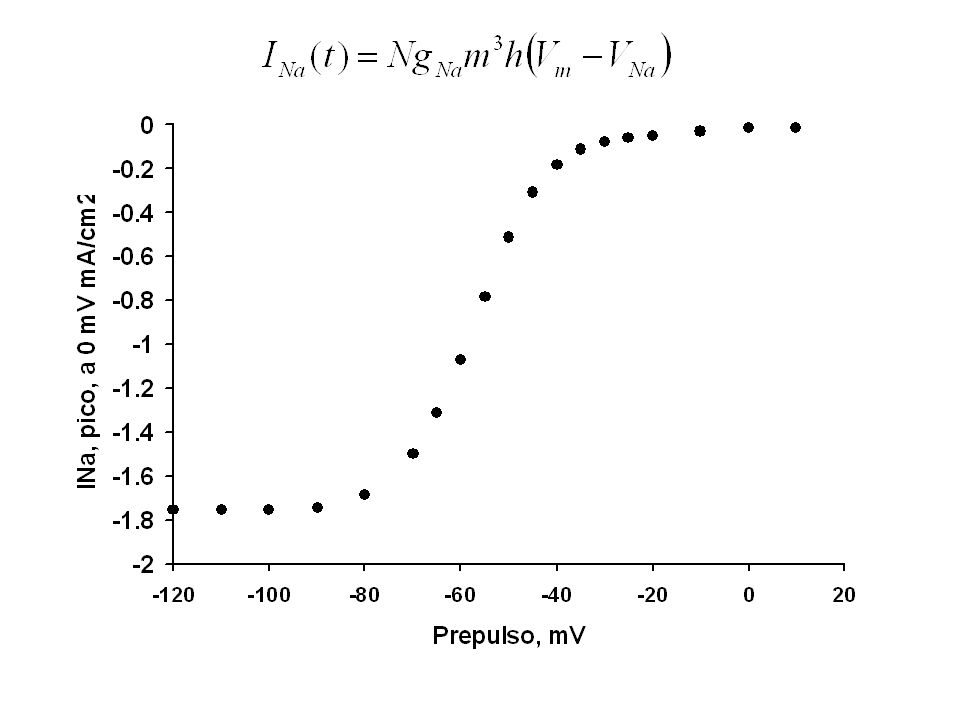
Canales de sodio del axón de jibia

62
La corriente de Na es la diferencia de la corriente control – corriente con TTX.
-60,8mV 0mV t, ms I, mA/cm2

63
Con el uso de TTX se puede aislar la fracción de la corriente llevada por los canales de sodio.
La corriente de Na presenta inactivación Vm 0 mV -60 mV

64
Procedimiento para demostrar que los canales de sodio cumplen la ley de Ohm y encontrar el potencial de inversión de la corriente La corriente de Na presenta inactivación 30 mV Vm 0 mV -60 mV

65
Sorpresa: El valor encontrado, 41 mV es menos positivo que el esperado según Nernst (54 mV).
Una explicación posible: los canales de Na no son idealmente selectivos. Calcule la razón GNa/GK para estos canales VNa = 41.2 mV Los canales de sodio cumplen la ley de Ohm

66
El modelo de bola y cadena para la inactivación de los canales de sodio
Figure Ball-and-Chain Model for Channel Inactivation. The inactivation domain, or "ball" (red), is tethered to the channel by a flexible "chain" (green). In the closed state, the ball is located in the cytosol. Depolarization opens the channel and creates a negatively charged binding site for the positively charged ball near the mouth of the pore. Movement of the ball into this site inactivates the channel by occluding it. [After C. M. Armstrong and F. Bezanilla. J. Gen. Physiol. 70(1977):567.]
, is tethered to the channel by a flexible chain (green). In the closed state, the ball is located in the cytosol. Depolarization opens the channel and creates a negatively charged binding site for the positively charged ball near the mouth of the pore. Movement of the ball into this site inactivates the channel by occluding it. [After C. M. Armstrong and F. Bezanilla. J. Gen. Physiol. 70(1977):567.]")
67
Topología de un canal de sodio membrana
Inactivation Gate

68
Tratando los axones con pronasa se puede estudiar la activación sin interferencia de la inactivación
0 mV -60 mV

69
Procedimiento para determinar la conductacia de los canales.
0 mV -40 mV -60 mV

70
INa,max = mAcm-2 a -40 mV VNa = 41.2 mV GNa,max = 116 mS cm-2

71
La función pNa(V,) no se puede ajustar bien con una función de Boltzmann
 no se puede ajustar bien con una función de Boltzmann")
72
Cinética de la activación
Vm 0 mV -40 mV -60 mV

73
x = 1 Suma de cuadrados = 8.0 La activación de los canales de sodio tiene un curso temporal sigmoidal. En el caso de los canales de potasio lo simulamos usando una función exponencial elevada a la cuarta potencia.

74
x = 2 Suma de cuadrados = 0.14 La activación de los canales de sodio tiene un curso temporal sigmoidal. En el caso de los canales de potasio lo simulamos usando una función exponencial elevada a la cuarta potencia.

75
x = 3 Suma de cuadrados = 0.00 La activación de los canales de sodio tiene un curso temporal sigmoidal. Lo simulamos usando una función exponencial elevada a la tercera potencia.

76
Con buenas estimaciones de m inicial y final para cada transición se puede encontrar tau m usando Solver.

77
m (0 mV) = 0.27 ms Con buenas estimaciones de m inicial y final para cada transición se puede encontrar tau m usando Solver.
 = 0.27 ms Con buenas estimaciones de m inicial y final para cada transición se puede encontrar tau m usando Solver.")
78
m (0 mV) = 0.27 ms m (-40mV) = 0.48 ms Con buenas estimaciones de m inicial y final para cada transición se puede encontrar tau m usando Solver.

79
Tau m en función del voltaje
La curva n vs Vm no se ajusta bien a una función secante hiperbólica.

80
Como ya sabemos la relación entre pNa y m podemos calcular de m(∞)
")
81
m() La curva m parece que se ajusta bien a una función de Boltzmann.
 La curva m parece que se ajusta bien a una función de Boltzmann.")
82
Como ya sabemos m y tau podemos calular alfa y beta

83
m y m a diferentes voltajes
Calcule m para el límite Vm -35 mV Ecuaciones empíricas de Hodkin y Huxley Ver “El impulso nervioso” por F. Bezanilla en libro Biofísica y Fisiología Celular. Eds. R.Latorre et al.

84
La inactivación. La variable h representa probabilidad de que los canales NO estén inactivados. Para h = 1 los canales no están inactivados. Para h = 0 los canales están inactivados La probabilidad de encontrar un canal conduciendo es m3h

85
Procedimiento para determinar h infinito en función del voltaje-
Medir la corriente en el mínimo. 0 mV -60 mV Potencial de mantención variable Pulso de prueba fijo en 0 mV

87
h() La curva h parece que se ajusta bien a una función de Boltzmann.
 La curva h parece que se ajusta bien a una función de Boltzmann.")
88
Determinación de h mediante ajuste de curvas al curso temporal de la corriente de sodio
-60 mV -60 mV -40 mV -100 mV

89
-60 mV -60 mV -40 mV -100 mV

90
h = 3.4 ms

91
Procedimiento para determinar h midiendo el curso temporal del escape de la inactivación
0 mV 0 mV -60 mV -60 mV

92
Procedimiento para determinar h midiendo el curso temporal del escape de la inactivación

93
Procedimiento para determinar h midiendo el curso tempooral del escape de la inactivación
h = 8.2 ms

94
Tau h en función del voltaje
La curva h vs Vm no se ajusta bien a una función secante hiperbólica.

95
Como ya sabemos m y tau en función de Vm, podemos calcular alfa y beta

96
Ecuaciones empíricas de Hodkin y Huxley 1952
Ecuaciones empíricas de Hodkin y Huxley Ver “El impulso nervioso” por F. Bezanilla en libro Biofísica y Fisiología Celular. Eds. R.Latorre et al.

97
h, ms h m, ms m NNagNa = 120 mS/cm2 VNa = 41,1 mV Vm, mV Vm, mV

98
Calcular la fracción de canales de sodio inactivados si se mantiene la membrana por mucho tiempo a -60 m. Calcular la fracción de canales de sodio abiertos después de 0.6 ms de despolarizar desde -60 mV a 0 mV. Calcular la fracción de canales de sodio inactivados después de 0.6 ms de despolarizar desde -60 mV a 0 mV. Calcular la fracción de canales de sodio cerrados después de 0.6 ms de despolarizar desde -60 mV a 0 mV.

99
Potencial de reposo, segunda visita

100
IK INa Im Vm IL IC En el potencial de reposo Im = 0 e IC = 0

101
IK INa Im Vm IL IC La tarea ahora es buscar un Vr se satisfaga esta ecuación. El problema lo puedo resolver porque conozco n, m y h en función de Vm.

102
Celda a ajustar para que la celda objetivo sea cero
h Celda Objetivo n Solver: Buscar que potencial hace que la intensidad de la corriente sea cero. m Vm ,(mV) Mapshow2010.xls
 Mapshow2010.xls.")
103
Solver: Este es el Potencial de Reposo.
Buscar que potencial hace que la intensidad de la corriente sea cero.

104
El potencial de acción de membrana

105
Computación de la trayectoria
Vm, n, m y h son los valores iniciales de la trayectoria. Con estas corrientes se calcula la derivada dVm/dt Ahora calculo el nuevo Vm Ahora calculo los nuevos valores de n, m y h para en cambio de voltaje desde Vm a Vm(t) por t unidades de tiempo.
 por t unidades de tiempo.")
106
Computación de la trayectoria
Con estos nuevos valores de Vm, n, m y h y las ecuaciones de las corrientes calculo la derivada dVm/dt Ahora calculo el nuevo Vm Ahora calculo los nuevos valores de n, m y h para en cambio de voltaje desde Vm a Vm(t) por t unidades de tiempo. Repito estos pasos para calcular toda la trayectoria de Vm(t)... Mostar Mapshow
 por t unidades de tiempo. Repito estos pasos para calcular toda la trayectoria de Vm(t)... Mostar Mapshow.")
107
Fin de la clase de canales de potasio del axón de jibia.
19 de Abril de 2010

Presentaciones similares