Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Estructura de proteínas, fuerzas que gobiernan el plegamiento
De la estructura unidimensional a la estructura tridimensional ● proteínas nativas y proteínas desnaturalizadas ● relaciones estructura-actividad La información para el plegamiento está contenida en la estructura primaria. Las proteínas nativas son estables? ● Christian Anfinsen y la ribonucleasa A Fuerzas que determinan el plegamiento (desde adentro), con una quimotripsina ● enlaces disulfuro ● enlaces iónicos ● enlaces de hidrógeno ● fuerzas de van der Waals Fuerzas que determinan el plegamiento (desde afuera) ● el agua y otros detalles El problema de decidir la estructura correcta ● ¿cuántos disulfuros puedes formar? ● la corta vida de una proteína Para ganar la carrera ● los pequeños motivos de una proteína ● un poco de ayuda: enzimas y chaperonas moleculares 7. Resumen
, con una quimotripsina. ● enlaces disulfuro. ● enlaces iónicos. ● enlaces de hidrógeno. ● fuerzas de van der Waals. Fuerzas que determinan el plegamiento (desde afuera) ● el agua y otros detalles. El problema de decidir la estructura correcta. ● ¿cuántos disulfuros puedes formar ● la corta vida de una proteína. Para ganar la carrera. ● los pequeños motivos de una proteína. ● un poco de ayuda: enzimas y chaperonas moleculares. 7. Resumen.")
2
Bibliografía Anfinsen, C and Scheraga, H. Experimental and theoretical aspects of protein folding. Adv. Prot. Chem 29, 205. Edelhoch, H. and Osborne, J. Thermodynamic Stability of Macromolecules. Adv. Prot. Chem.; 30; 200. Rossmann, M. and Argos, P. Protein Folding. Ann Rev. Biochem. 50; 497. Dressler, D. And Potter, H.; Discovering Enzymes; Scientific American Library, Vol 34. Chap 4. Kolata, G. Trying to crack the second half of the genetic code; Science, 233; 1039. Dobson, C. Evans, P, Radford, S, Understanding how proteins Fold: The Lysozyme Story so far, TIBS, 19, 31 Hartl, F. Hlodan, R. and Langer, T., Molecular Chaperones in protein folding: The art of avoiding Sticky situations., TIBS 19, 20 Hartl, F. and Hayer-Hartl, M. Converging concepts of protein folding in vitro and in vivo. Nature Structural & Molecular Biology 16: 574

3
Las proteínas se pliegan en forma específica y rápida in vivo apenas son sintetizadas

4
¿Son estables las Proteínas nativas?

5
¿Son estables las proteínas nativas?
¿qué quiere decir estable? ¿Qué quiere decir estable? ¿Qué consecuencias tendría una u otra hipótesis?

6
La información para el plegamiento está contenida en la estructura primaria
Christian Anfinsen y la ribonucleasa A

7
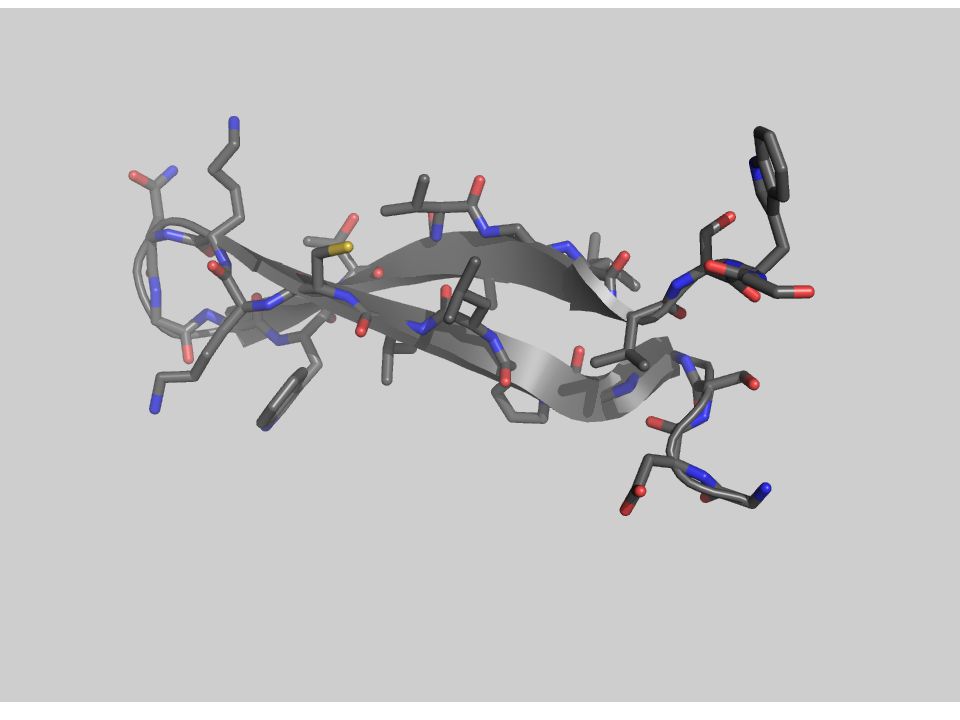
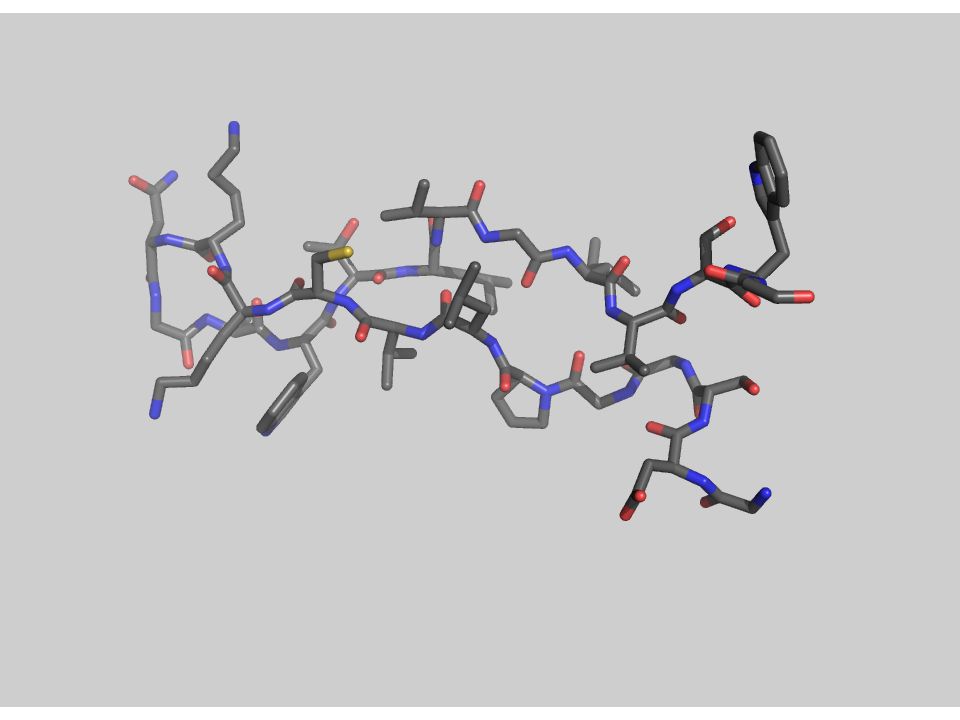
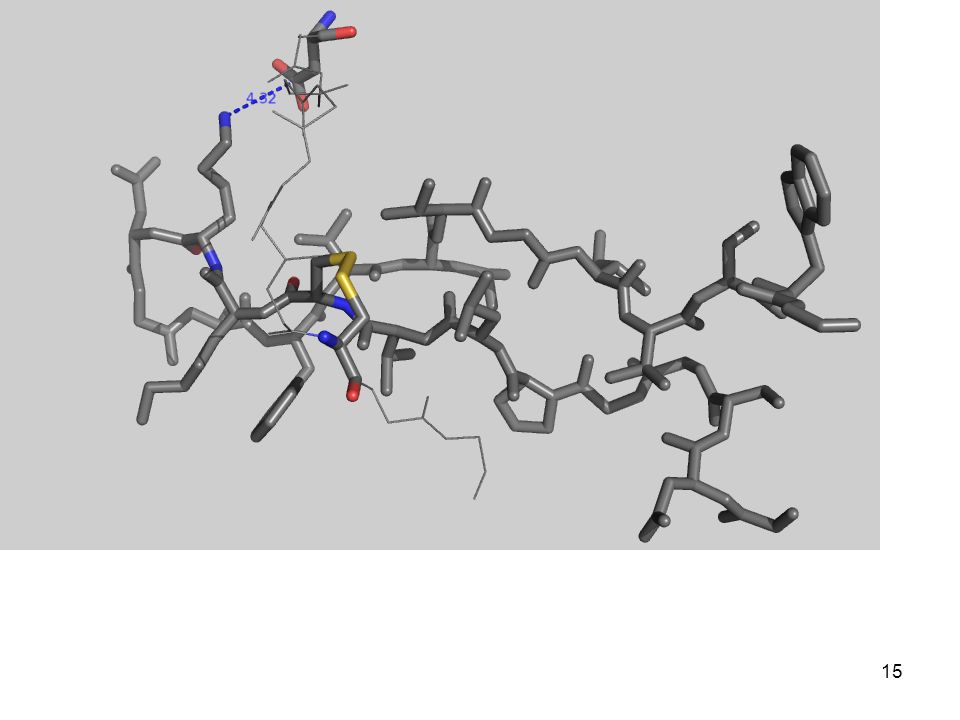
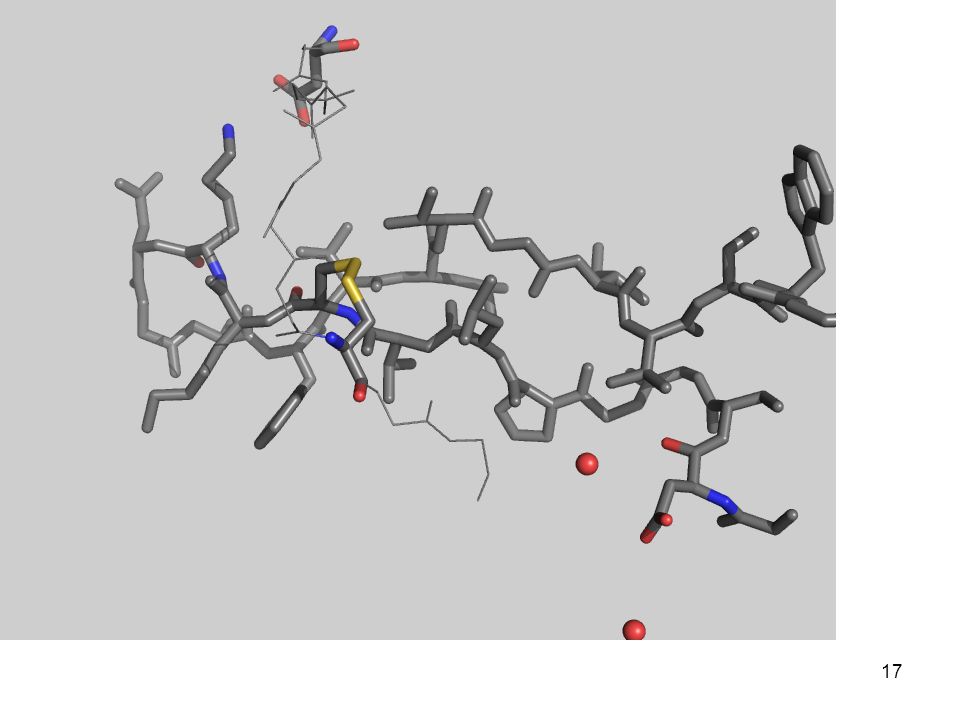
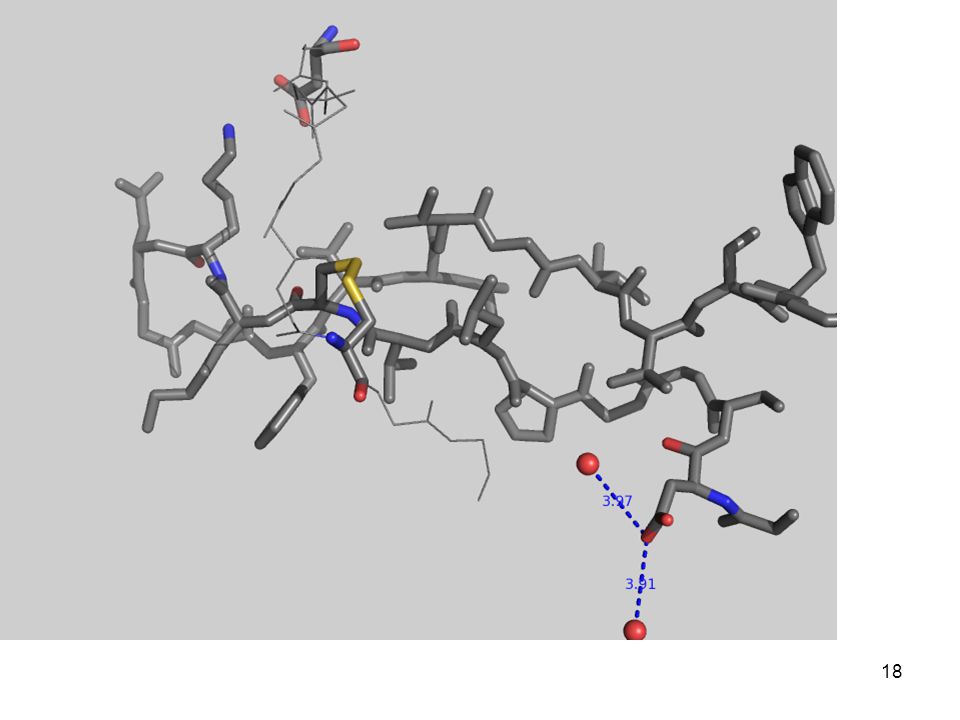
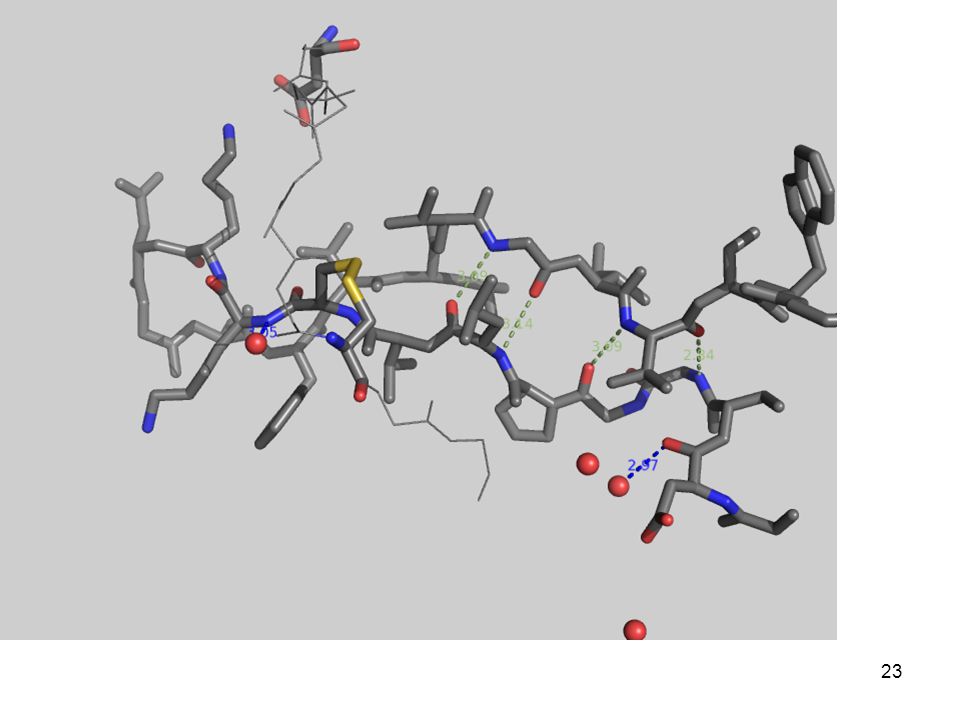
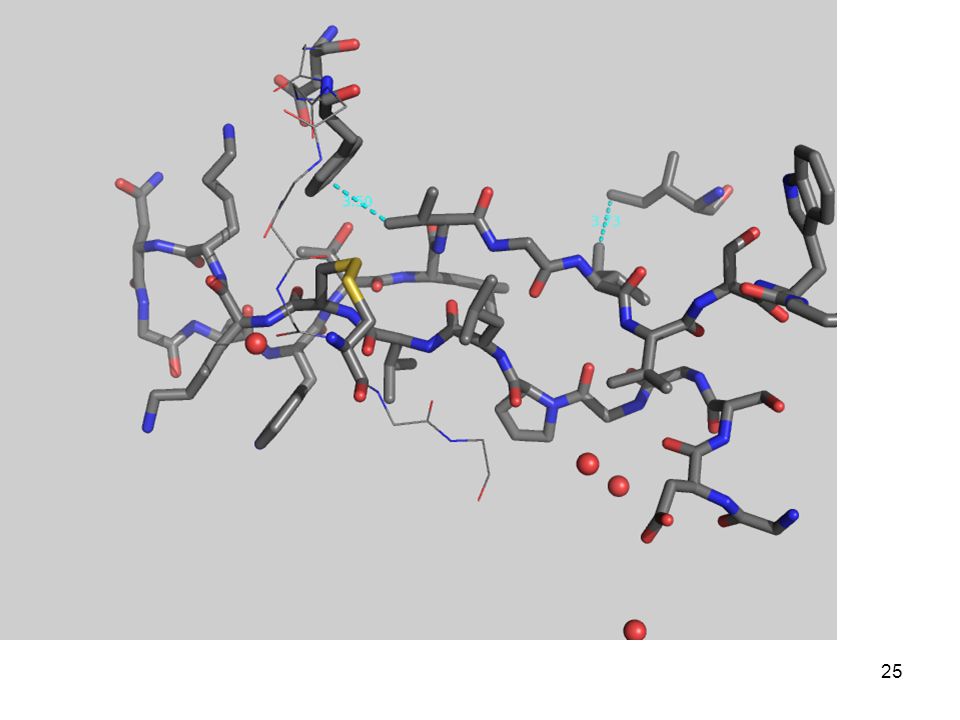
3.- Fuerzas que determinan el plegamiento
(desde adentro) Con una quimotripsina Péptido modelo: Gly-Asp-Ser-Gly-Gly-Pro-Leu-Val-Cys-Lys-Lys-Asn-Gly-Ala-Trp-Thr-Leu-Val-Gly-Ile-Val-Ser Trp Aminoácidos
 Con una quimotripsina. Péptido modelo: Gly-Asp-Ser-Gly-Gly-Pro-Leu-Val-Cys-Lys-Lys-Asn-Gly-Ala-Trp-Thr-Leu-Val-Gly-Ile-Val-Ser Trp. Aminoácidos")
11
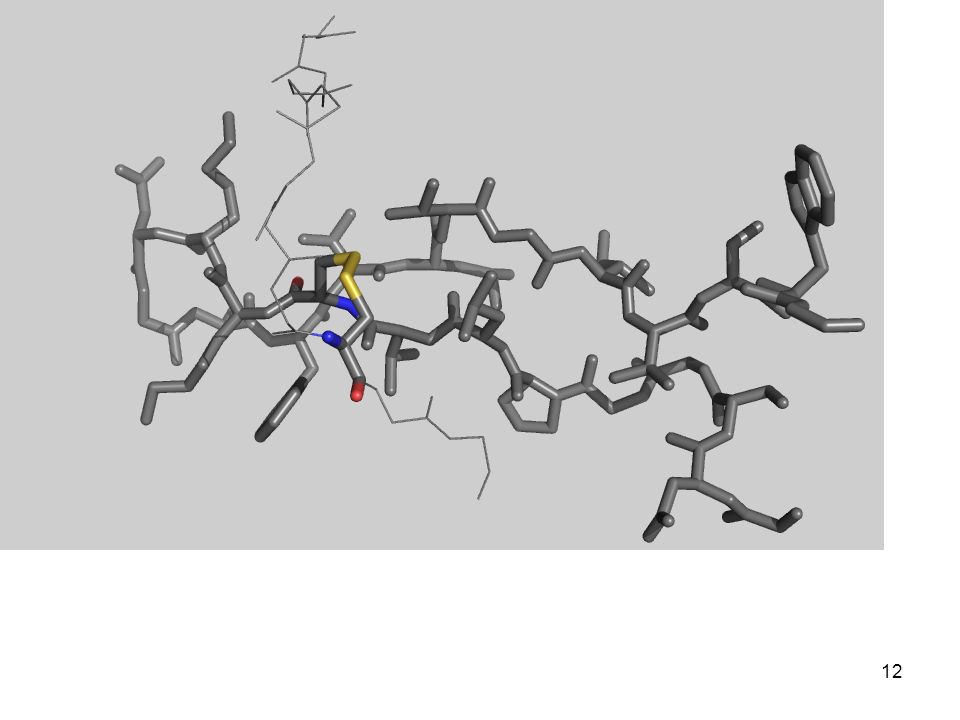
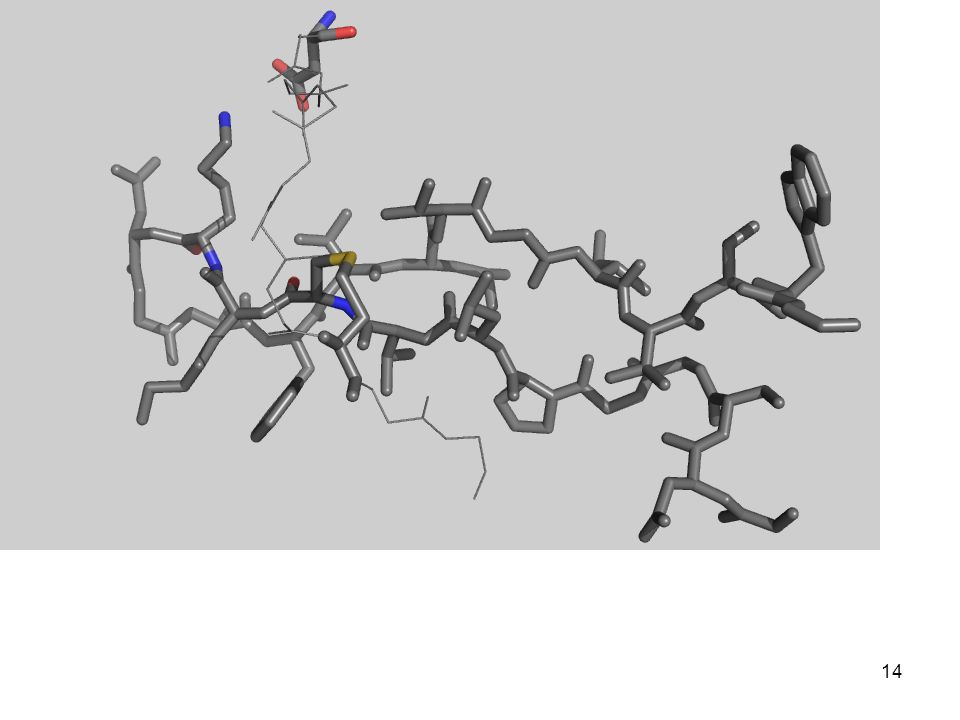
Interacciones covalentes Enlace disulfuro
Es un enlace covalente Involucra a dos residuos de cisteína Implica el intercambio de dos electrones (es decir, su formación o ruptura es una reacción redox) 2 RSH + Oxidante RS—SR + Producto RS—SR + Reductor RSH + Producto
 2 RSH + Oxidante RS—SR + Producto. RS—SR + Reductor 2 RSH + Producto.")
13
+ - Interacciones de carga Ion-Ion; Enlace iónico; puente salino
Lisina Arginina Amino terminal (Histidina) - Aspartato Glutamato Carboxilo terminal (Cisteína) = constante dieléctrica - vacío: e = 1 - medio apolar: e = 3-5 - agua: e = 78
 - Aspartato. Glutamato. Carboxilo terminal. (Cisteína) = constante dieléctrica. - vacío: e = 1. - medio apolar: e = agua: e = 78.")
16
Interacciones con dipolos
Ion dipolo d- d+ el dipolo se orienta y se atrae con una energía:

19
Interacciones con dipolos
Dipolo dipolo d- d+ d- d+ Los dipolos se ORIENTAN y se atraen con una energía: pero además los dipolos se SUMAN

20
d- d+

21
Interacciones con dipolos
Enlace de hidrógeno. Básicamente es una interacción dipolo-dipolo con un átomo de hidrógeno en el medio d- d+ d- d+ Altamente DIRECCIONAL donador receptor 2.7 Å E = 1-3 kcal/mol

22
Interacciones con dipolos
Enlace de hidrógeno. Un enlace de hidrógeno entre un receptor y un donador tiene múltiples configuraciones posibles, por ejemplo Serina … y todas tienen prácticamente la misma energía Asparagina

24
Interacciones de van der Waals
Son interacciones sumamente DÉBILES entre moléculas. Implican dipolos instantáneos y dipolos inducidos. Dependen (entre otras cosas) del número de electrones de la molécula (P.M.), por ejemplo: CH4 Metano (gas) C8H18 Nafta (líquido) C40H82 Parafina (sólido) Son interacciones de contacto (E α r-6). Están presentes en TODOS LOS CASOS.
 del número de electrones de la molécula (P.M.), por ejemplo: CH4 Metano (gas) C8H18 Nafta (líquido) C40H82 Parafina (sólido) Son interacciones de contacto (E α r-6). Están presentes en TODOS LOS CASOS.")
26
Resumen de interacciones
Por supuesto que estos son valores promedio y aproximados. Pero destacan la importancia del AGUA Tipo de enlace Energía (kcal/mol) Distancia (Å) En el vacío En agua Covalente 1.5 90 Iónico 2.5 80-200 ~ 3 Hidrógeno 3.0 4 1 – 3 van der Waals 3.5 0.1
 Distancia (Å) En el vacío. En agua. Covalente Iónico ~ 3. Hidrógeno – 3. van der Waals")
27
Los solutos no polares odian al agua por eso se van Versión 2
4. Fuerzas que determinan el plegamiento (desde afuera) El agua y otros detalles (efecto hidrofóbico) ¿Quién odia al agua? Describamos el fenómeno: Un soluto no polar en agua tiende a precipitar, agruparse, huir, etc. Versión 1 Los solutos no polares odian al agua por eso se van Versión 2 El agua expulsa a los solutos no polares porque prefiere estar sola
 El agua y otros detalles (efecto hidrofóbico) ¿Quién odia al agua Describamos el fenómeno: Un soluto no polar en agua tiende a precipitar, agruparse, huir, etc. Versión 1. Los solutos no polares odian al agua por eso se van. Versión 2. El agua expulsa a los solutos no polares porque prefiere estar sola.")
28
¿Hay simpatías entre moléculas?
4. Fuerzas que determinan el plegamiento (desde afuera) ¿Hay simpatías entre moléculas? DG = DH -TDS El agua y los solutos van a hacer todo lo posible para que: DH < 0 DS > 0 DG < 0 El efecto hidrofóbico puede entenderse en términos de la energía de Gibbs del sistema
 ¿Hay simpatías entre moléculas DG = DH -TDS. El agua y los solutos van a hacer todo lo posible para que: DH < 0. DS > 0. DG < 0. El efecto hidrofóbico puede entenderse en términos de la energía de Gibbs del sistema.")
29
+ DG < 0 4. Fuerzas que determinan el plegamiento (desde afuera)
H disminuye (se forman enlaces) S aumenta (se liberan moléculas) DG < 0
 S aumenta (se liberan moléculas) DG < 0.")
30
4. Fuerzas que determinan el plegamiento (desde afuera)
Simpatías entre moléculas Escalas de hidropatías para cadenas laterales de aminoácidos Kyte y Doolittle (1982) Arg Lys Asn Asp Gln Glu His Pro Tyr Trp -4.5 -3.9 -3.5 -3.2 -1.6 -1.3 -0.9 Ser Thr Gly Ala Met Cys Phe Leu Val Ile -0.8 -0.7 -0.4 1.8 1.9 2.5 2.8 3.8 4.2 4.5
 Arg. Lys. Asn. Asp. Gln. Glu. His. Pro. Tyr. Trp Ser. Thr. Gly. Ala. Met. Cys. Phe. Leu. Val. Ile")
31
4. Fuerzas que determinan el plegamiento (desde afuera)
Esta escala no permite predecir estructuras tridimensionales pero da indicios de algunas características mediante mapas de hidropatía Albúmina

32
4. Fuerzas que determinan el plegamiento (desde afuera)
Esta escala no permite predecir estructuras tridimensionales pero da indicios de algunas características mediante mapas de hidropatía Hemoglobina

33
4. Fuerzas que determinan el plegamiento (desde afuera)
Esta escala no permite predecir estructuras tridimensionales pero da indicios de algunas características mediante mapas de hidropatía Citocromo oxidasa

34
5. El problema de decidir la estructura correcta
¿Cuántos disulfuros se pueden formar? Cisteínas Disulfuros posibles Combinaciones posibles 2 1 4 3 6 15 8 105 10 5 945 12 10395 14 7 135135 16 34 17 46 23 ← RNAsa A ← Quimotripsina ← Albúmina sérica ← Inmunoglobulina Edad del universo (s)
 ")
35
? 5. El problema de decidir la estructura correcta
El plegamiento no puede ser un proceso de búsqueda aleatoria. Debe existir un camino secuencial y ordenado para llegar al estado nativo. El “camino” debe proporcionar una ruta RÁPIDA e INEQUÍVOCA hacia el estado nativo. Además, la ruta debe estar codificada en la secuencia de aminoácidos ya que muchas proteínas se pliegan espontáneamente in vitro Secuencia de aminoácidos ? Estructura 3D alguna ayudita

36
6. Para ganar la carrera Los pequeños motivos de una proteína
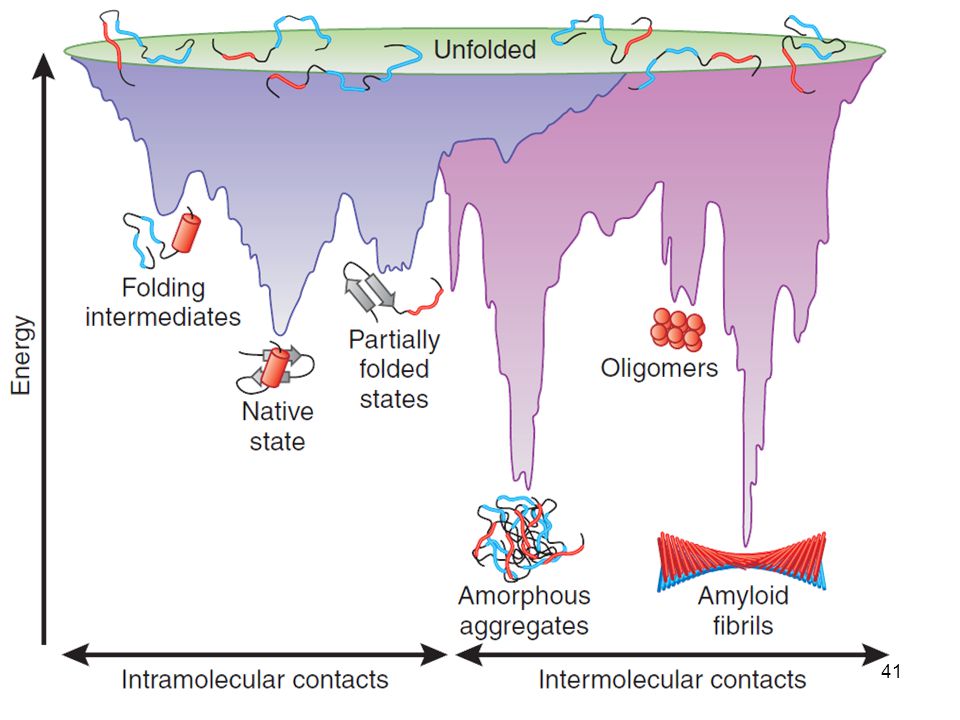
IN VITRO: El plegamiento de un polipéptido enrollado al azar comienza con la formación de pequeñas secciones de estructura secundaria, como hélices a, giros b, etc., que pueden funcionar como núcleos desde donde el plegamiento se extiende. Los núcleos que tienen la estructura adecuada pueden crecer por la difusión, colisión o adhesión de dos o más núcleos, hasta formar los dominios de la proteína Muchas proteínas se pliegan a su estado nativo en una secuencia ordenada de pequeños “colapsos” que forman “motivos” de la estructura 3D.

37
6. Para ganar la carrera En proteínas con más de un dominio, estos se pueden juntar para formar un “glóbulo fundido” que tiene la estructura secundaria correcta pero tiene una estructura terciaria desordenada, por ejemplo, puede haber residuos hidrofóbicos expuestos al disolvente.

38
6. Para ganar la carrera Un poco de ayuda
Isomerasas de disulfuros proteicos Isomerasas de prolinas peptídicas cis-trans El plegamiento de las proteínas tiene algunos pasos más lentos como la formación de enlaces disulfuro o la isomerización cis-trans de los enlaces peptídicos de prolina Para acelerar estos procesos hay, por supuesto, enzimas

39
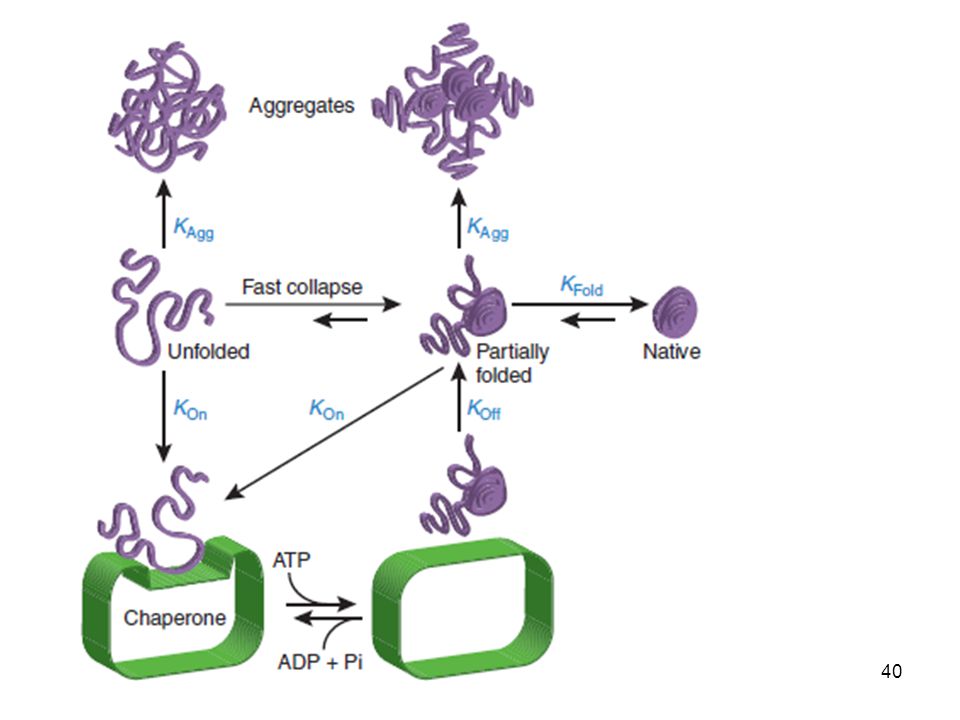
6. Para ganar la carrera Un poco de ayuda Chaperonas moleculares
Proteínas de shock térmico Las chaperonas moleculares inhiben interacciones inapropiadas entre superficies potencialmente complementarias y rompen uniones inadecuadas para facilitar asociaciones más funcionales Las Hsp se unen a cadenas polipeptídicas nacientes cuando van emergiendo del ribosoma. Revierten la desnaturalización y la agregación de proteínas

42
Netzer, WJ and Hartl, FU; TIBS, (1998) 23: 68
 23: 68")
43
Estructura de proteínas, fuerzas que gobiernan el plegamiento
De la estructura unidimensional a la estructura tridimensional ● proteínas nativas y proteínas desnaturalizadas ● relaciones estructura-actividad La información para el plegamiento está contenida en la estructura primaria. Las proteínas nativas son estables? ● Christian Anfinsen y la ribonucleasa A Fuerzas que determinan el plegamiento (desde adentro), con una quimotripsina ● enlaces disulfuro ● enlaces iónicos ● enlaces de hidrógeno ● fuerzas de van der Waals Fuerzas que determinan el plegamiento (desde afuera) ● el agua y otros detalles El problema de decidir la estructura correcta ● ¿cuántos disulfuros puedes formar? ● la corta vida de una proteína Para ganar la carrera ● los pequeños motivos de una proteína ● un poco de ayuda: enzimas y chaperonas moleculares 7. Resumen
, con una quimotripsina. ● enlaces disulfuro. ● enlaces iónicos. ● enlaces de hidrógeno. ● fuerzas de van der Waals. Fuerzas que determinan el plegamiento (desde afuera) ● el agua y otros detalles. El problema de decidir la estructura correcta. ● ¿cuántos disulfuros puedes formar ● la corta vida de una proteína. Para ganar la carrera. ● los pequeños motivos de una proteína. ● un poco de ayuda: enzimas y chaperonas moleculares. 7. Resumen.")
Presentaciones similares






 Noviembre de 2004.>")
 29 de julio de 2004.>")












