Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Sistema de Lanzaderas Cadena Respiratoria Fosforilación Oxidativa
Dra. Ingrid Estévez

2
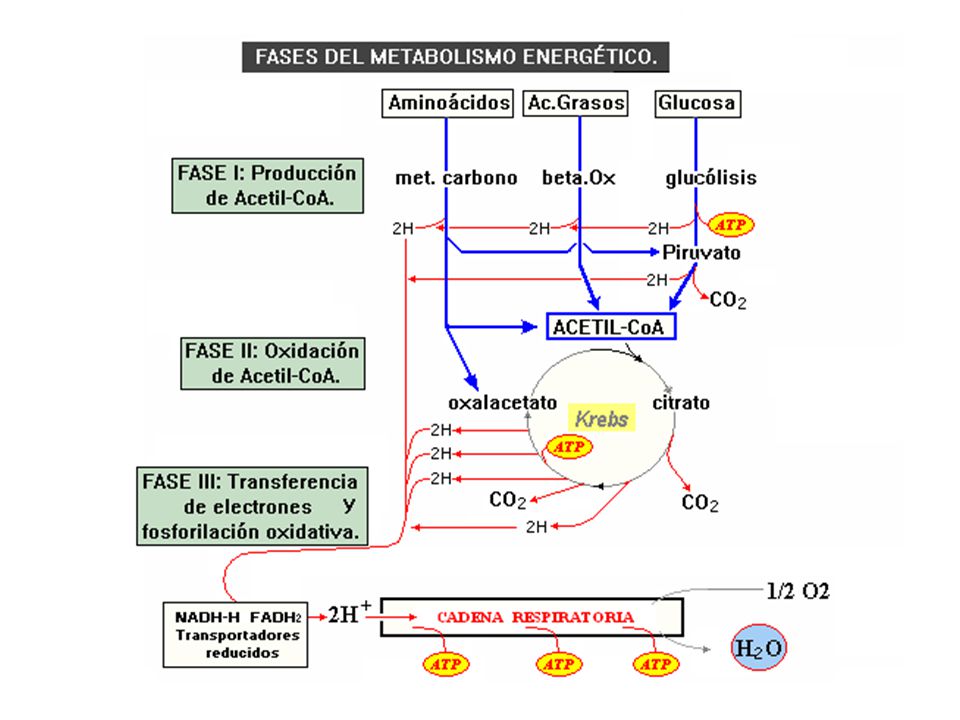
MITOCONDRIA PRINCIPAL FUNCIÓN: OXIDACIÓN DE COMBUSTIBLES METABÓLICOS Y CONSERVA ENERGÍA LIBRE PARA SINTESIS DE ATP

4
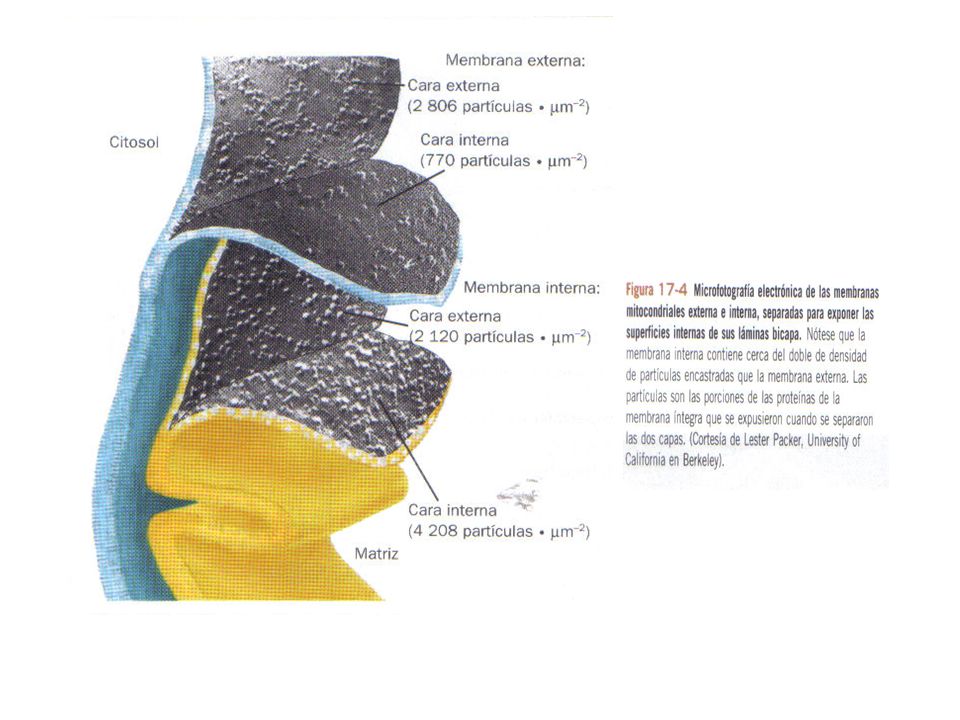
Mitocondria Membrana externa: Membrana interna:
Enzimas y proteínas de transporte (porinas) Permeable a moléculas pequeñas, iones y proteínas de menos de D Membrana interna: Impermeable a la mayoría de las moléculas Contiene: Transportadores de electrones (I- IV) ADP – ATP translocasas ATP sintasa
 Permeable a moléculas pequeñas, iones y proteínas de menos de D. Membrana interna: Impermeable a la mayoría de las moléculas. Contiene: Transportadores de electrones (I- IV) ADP – ATP translocasas. ATP sintasa.")
5
Mitocondria Matriz: Contiene: Complejo Piruvato deshidrogenasa
Enzimas ciclo de Krebs Enzimas b-oxidación Enzimas oxidación de aminoácidos DNA, ribosomas ATP, ADP, Pi, Mg, Ca, K

6
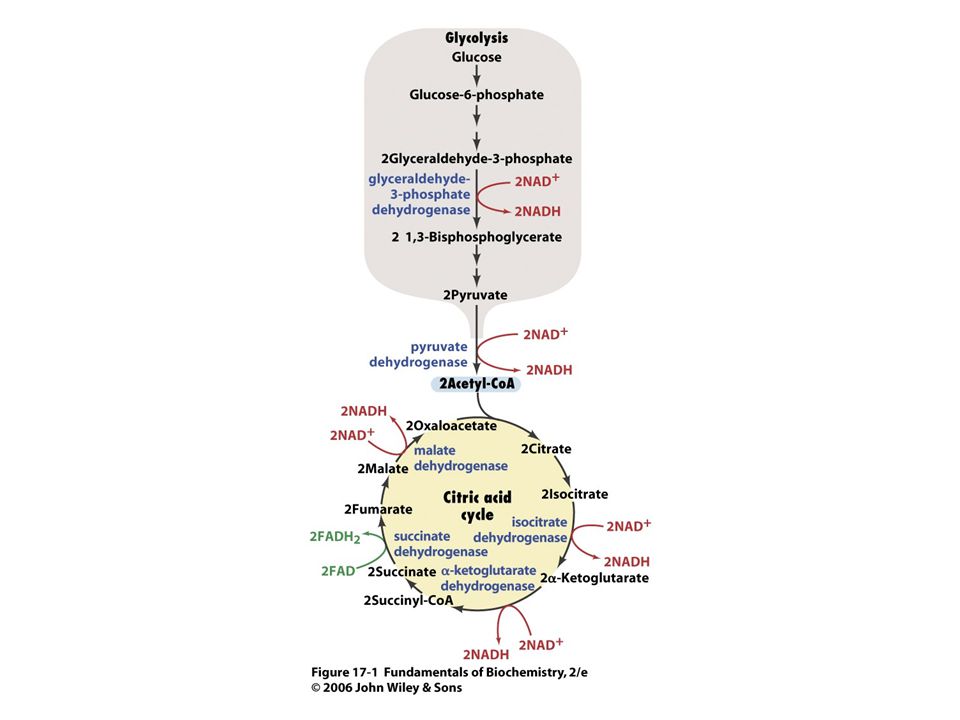
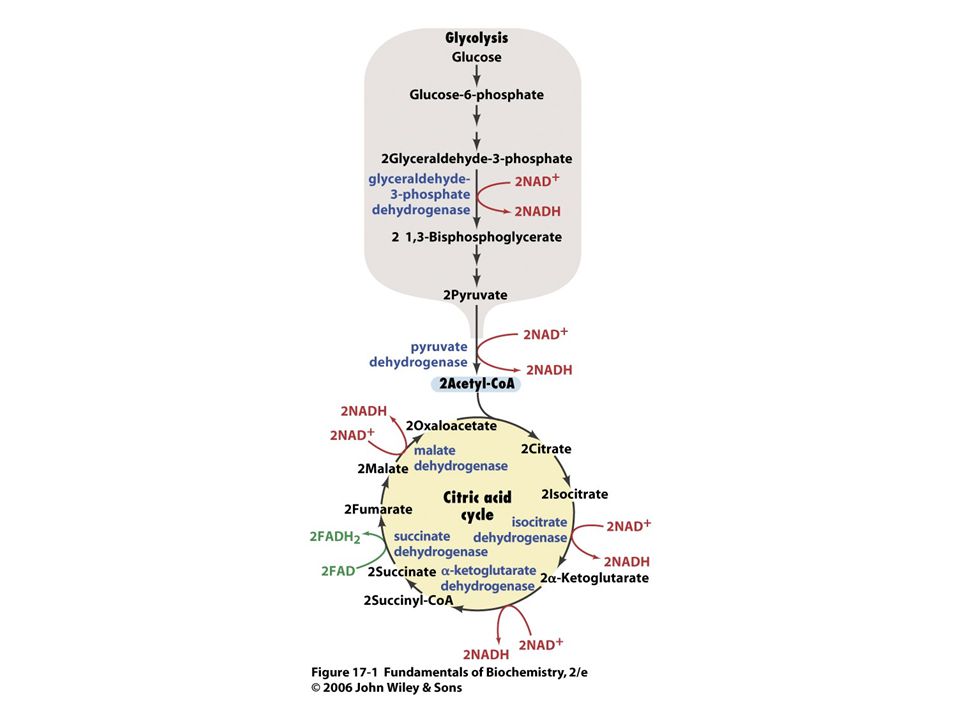
Origen del ATP Fosforilación a nivel de sustrato
Fosforilación Oxidativa (Fosforilación a nivel de Cadena Respiratoria).
.")
7
Fosfoglicerato Cinasa
La Fosforilación a nivel de sustrato es un mecanismo poco habitual de formación de ATP FOSFOGLICERATO CINASA Fosfoglicerato Cinasa 1,3 Bis-fosfoglicerato ADP Fosfoglicerato ATP SUCCINIL CoA sintetasa (Ciclo de Krebs)
")
8
Malato Deshidrogenasa Succinato Deshidrogenasa
NAD y FAD son los receptores de los electrones en las reacciones de las rutas metabólicas Malato Deshidrogenasa L- Malato Oxalacetato Succinato Deshidrogenasa Succinato Fumarato

10
LANZADERA DE GLICEROL FOSFATO

11
glucolisis Matriz Glicerol 3 fosfato Dihidroxiacetona fosfato
deshidrogenasa citosolica Glicerol 3 fosfato Dihidroxiacetona fosfato Glicerol 3 fosfato deshidrogenasa mitocondrial Matriz

12
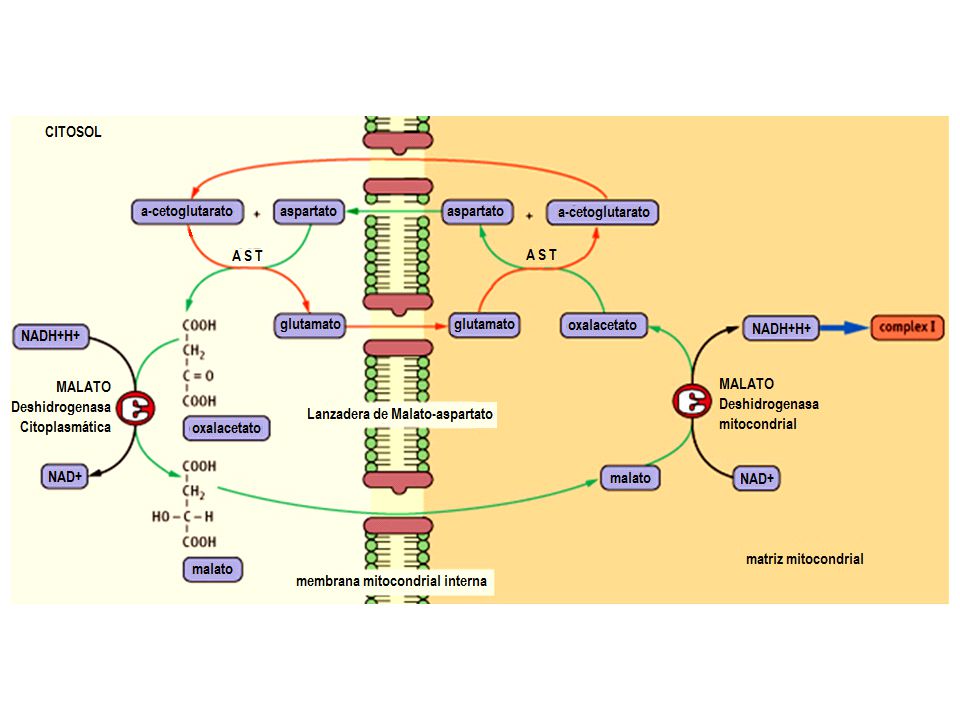
LANZADERA DE MALATO-ASPARTATO

15
Reaction moles ATP/mol Glc hexokinase phosphofructokinase G3PDH
Mechanism moles ATP/mol Glc hexokinase phosphorylation -1 phosphofructokinase G3PDH NADH, oxidative phosphorylation +6 (+4)* phosphoglycerate kinase substrate-level phosphorylation 2 pyruvate kinase pyruvate dehydrogenase 6 isocitrate dehydrogenase α-ketoglutarate dehydrogenase succinyl CoA synthetase substrate-level phosphorylation (GTP) succinate dehydrogenase FADH2, oxidative phosphorylation 4 malate dehydrogenase TOTAL 38 (36) Son 40 (38) ATP totales pero se le restan dos de la fase de inversión en la glucólisis. Y netos serán 36 si se pasan los equivalentes reductores usando la lanzadera de glicerol fosfato porque la coenzima mitocondrial será FADH2, mientras que si usa la lanzadera de malato-aspartato serán 38 netos porque la coenzima mitocondrial es NADH.
* phosphoglycerate kinase. substrate-level phosphorylation. 2. pyruvate kinase. pyruvate dehydrogenase. 6. isocitrate dehydrogenase. α-ketoglutarate dehydrogenase. succinyl CoA synthetase. substrate-level phosphorylation (GTP) succinate dehydrogenase. FADH2, oxidative phosphorylation. 4. malate dehydrogenase. TOTAL. 38 (36) Son 40 (38) ATP totales pero se le restan dos de la fase de inversión en la glucólisis. Y netos serán 36 si se pasan los equivalentes reductores usando la lanzadera de glicerol fosfato porque la coenzima mitocondrial será FADH2, mientras que si usa la lanzadera de malato-aspartato serán 38 netos porque la coenzima mitocondrial es NADH.")
17
Cadena Respiratoria y Fosforilación Oxidativa
Cadena Respiratoria: Transporte en secuencia ordenada de los equivalentes reducidos desde los sustratos donadores hasta la formación final de AGUA. Fosforilación Oxidativa: Sistema de conversión o captura de la energía liberada en la cadena respiratoria (68%), para unir ADP + Pi y formar ATP.
, para unir ADP + Pi y formar ATP.")
18
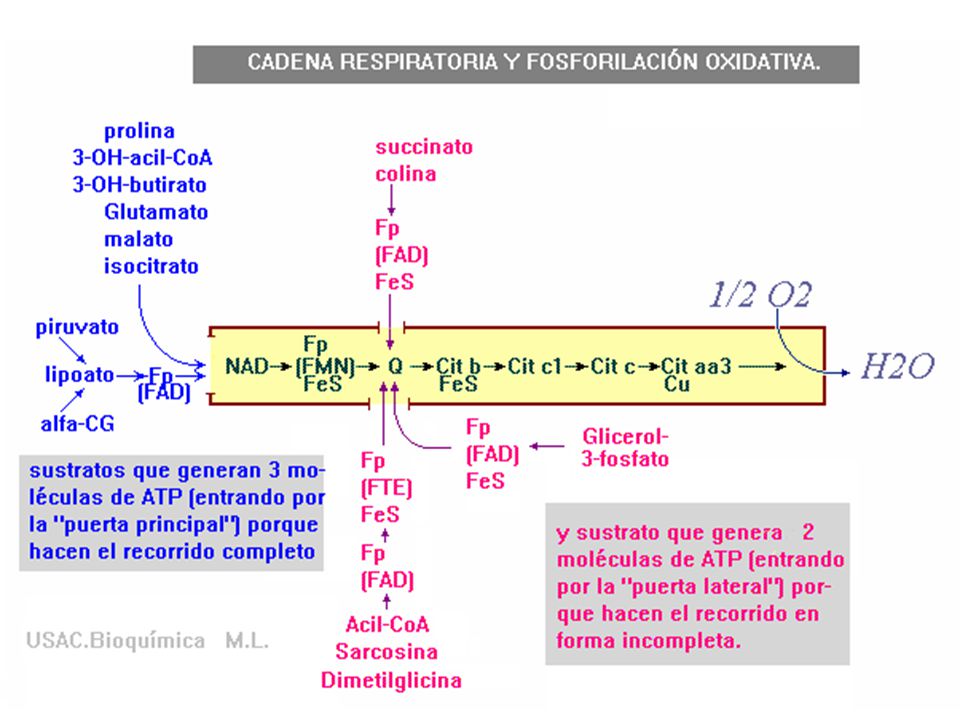
CADENA RESPIRATORIA Y FOSFORILACIÓN OXIDATIVA
Traslocación de protones

19
El hidrógeno y los electrones fluyen a lo largo de la cadena en etapas, a partir de los componentes de mayor potencial redox negativo hacia los componentes de mayor potencial redox positivo

21
CADENA RESPIRATORIA

22
Complejo I NADH-Q- Oxidorreductasa NADH DESHIDROGENASA
Constituido por: FMN, centros ferrosulfurados. Cataliza transferencia de un par de e- del NADH a la ubiquinona (UQ)
")
23
Complejo II Succinato-Q- Oxidoreductasa succinato deshidrogenasa
Compuesto por: FAD, Centro ferrosulfurada Transferencia de e- del succinato al FAD y a la ubiquinona.

24
El coenzima Q es paso obligatorio de los electrones procedentes de varias vías
Espacio intermembranoso Succinato Matriz Flavoproteína de transferencia de electrones

25
Complejo III Ubiquinona citocromo c oxidorreductasa o citocromo bc1
Constituido por: grupos hemo, ferrosulfurados Cataliza la transferencia de e- desde la ubiquinona reducida (UQH2) al citocromo bc1
 al citocromo bc1.")
26
CICLO Q (Complejo III) Oxidación de la Oxidación de la primera QH2
segunda QH2 Espacio Intermembrana Matriz

27
complejo IV citocromo oxidasa
Formado por grupos hemo (a,a3), ión cobre Paso final del transporte de e- Transferencia de e- del citocromo c al oxigeno formando 2 H2O
, ión cobre. Paso final del transporte de e- Transferencia de e- del citocromo c al oxigeno formando 2 H2O.")
28
Gradiente quimiosmótico
El transporte de electrones origina un gradiente de protones

29
TEORIA QUIMIOSMOTICA •Propuesta por Peter Mitchell en losaños60 (Premio Nobel 1978) •Teoría Quimiosmótica: Gradiente de protones electroquímico Este gradiente sirve para sintetizar ATP

31
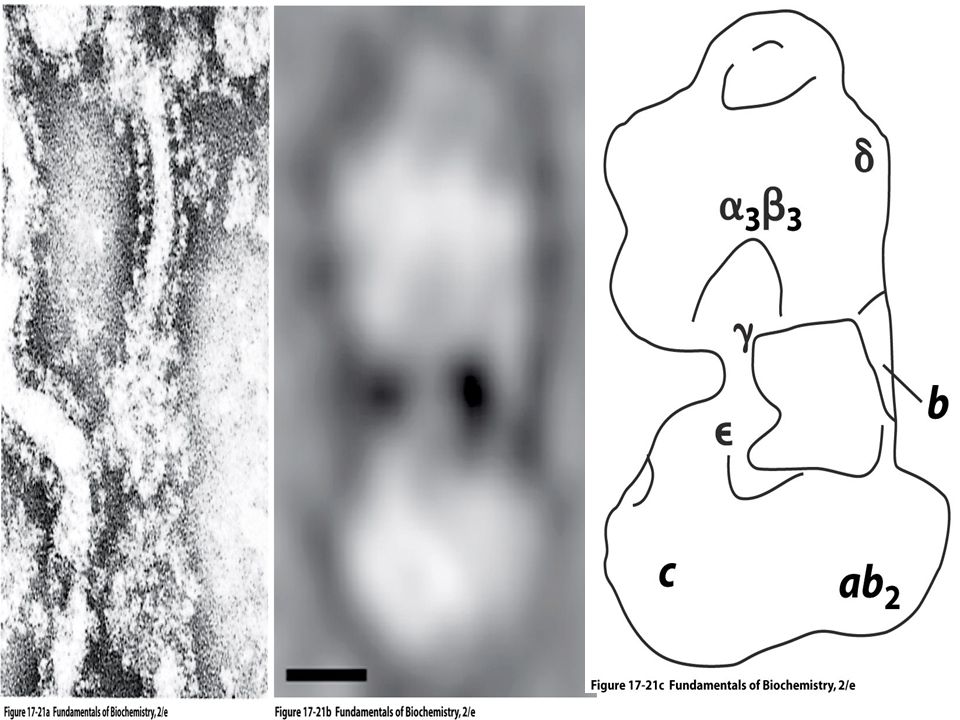
Complejo V ATP - SINTASA
Varias subunidades 2 elementos: F1: Factor 1 de acoplamiento Fo: Sección basal, integrada a la membrana interna, canal de protones

32
ATP - SINTASA F1: Fo: 3 subunidades a 3 subunidades b
Eje central subunidad g Fo: 3 subunidades (a, b, c)
")
33
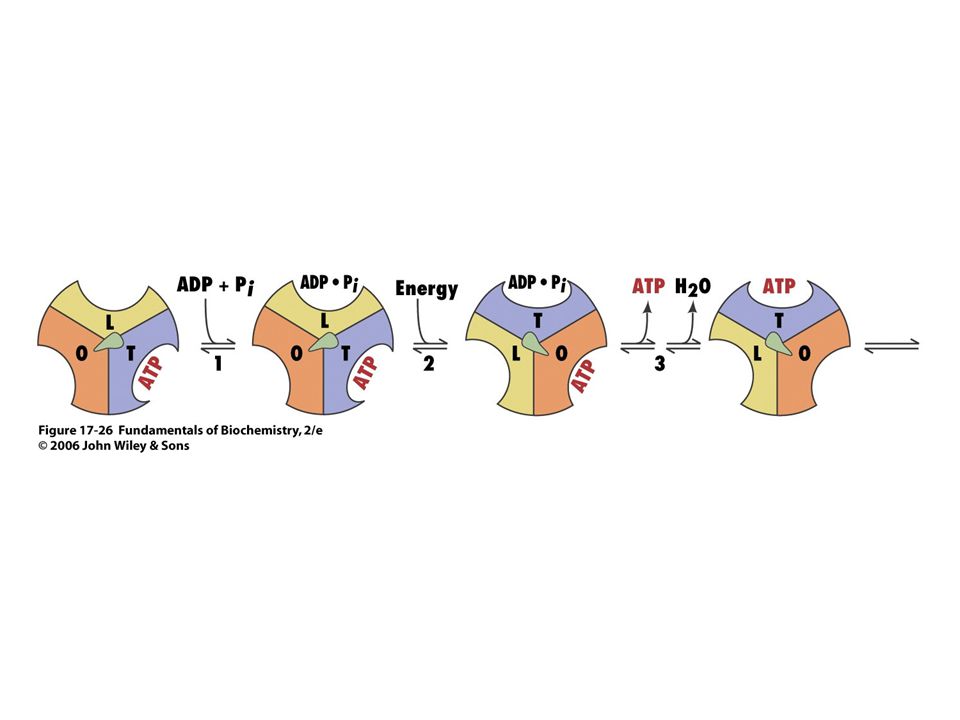
La ATP sintasa es un rotor
Filamento de Actina El anillo c gira con el paso de los H+. El giro de las subunidades ge provoca cambios conformacionales en las subunidades a b según va girando el complejo por el paso de 3 H+ Complejo Ni

34
•La rotación de γ es producida por el paso de protones
a través de la subunidades, que produce una rotación del anillo de subunidades C Rotación

35
•La Fosforilación Oxidativa es simple en cuanto a concepto, pero compleja en cuanto al mecanismo.
(1) Cadena de transporte electrónica Conjunto de complejos enzimáticos colocados en la membrana mitocondrial que oxidan NADH y FADH2 generándose un gradiente de protones (2) ATP sintasa aprovecha la energía del gradiente de protones para producir ATP
 Cadena de transporte electrónica Conjunto de complejos enzimáticos colocados en la membrana mitocondrial que oxidan NADH y FADH2 generándose un gradiente de protones. (2) ATP sintasa aprovecha la energía del gradiente de protones para producir ATP.")
36
F0: bomba de H+ •Segmento hidrofóbico que atraviesa la membrana interna mitocondrial •Contiene conducto de H + del complejo •Formado por subunidades c -1 subunidad a en la periferia del anillo Matriz M. Espacio Intemembrana

37
•F1 y F0 estanconectadas por el tallo γεy por una columna externa formada por b2 y δ
•Podemos considerar que el enzima esta formado por dos componentes funcionales: -UNIDAD MOVIL: anillo c y tallo γ ε -UNIDAD ESTATICA: resto de la molécula Matriz M. Espacio Intemembrana

38
Mecanismo de rotación del anillo c
•cada protón entra por el semiconducto citosólico, sigue una vuelta completa por el anillo c y sale por el otro semiconducto hacia la matriz •Según este modelo: el numero de protones que se han de transportar para generar una molécula de ATP dependerá del número de subunidades del anillo c• .Si el anillo tiene 10 subunidades(ATP sintasa de levadura): cada vuelta del anillo generara 3 ATP y fluiran10 protones: 10/3 ~ 3 H + por ATP
: cada vuelta del anillo generara 3 ATP y fluiran10 protones: 10/3 ~ 3 H + por ATP.")
41
BALANCE ENERGÉTICO ORIGEN DEL ATP
Proceso Citoplasma Matriz Mitocondrial Transporte Electrónico GLUCOLISIS 2 ATP 2 NADH Sistema de Lazaderas 2 x (ATP) 3 x (2NADH) 6 ATP Piruvato a Acetil CoA Desc. O. Piruvato RESPIRACION Ciclo de Krebs 2 X (NADH +H) 2 X (ATP) 2 X (3 NADH + H) 2 X (1 FADH) 2 X (3ATP) 6 X (3 ATP) 2 X (2 ATP) 18 ATP 4 ATP 36 ATP BALANCE ENERGÉTICO (POR CADA MOLECULA DE GLUCOSA ATP
 3 x (2NADH) 6 ATP. Piruvato. a Acetil CoA. Desc. O. Piruvato. RESPIRACION. Ciclo de Krebs. 2 X (NADH +H) 2 X (ATP) 2 X (3 NADH + H) 2 X (1 FADH) 2 X (3ATP) 6 X (3 ATP) 2 X (2 ATP) 18 ATP. 4 ATP. 36 ATP. BALANCE ENERGÉTICO (POR CADA MOLECULA DE GLUCOSA 38 ATP.")
42
Inhibidores del Transporte Electrónico
FADH Rotenona Antimicina A CN-,CO, H2S

43
Antimicina Tenoil Trifluro acetato DCCD Oligomicina
Gradiente de Protones Monoxido de C Acido Sulfridico Cianuro Antimicina Complejo I Complejo III Complejo II Succinato Piericidina A Amobarbital Rotenona Mercuriales Esteroides Tenoil Trifluro acetato NADH DCCD Oligomicina Complejo IV Desacopladores: 2-4 dinitrofenol, Dicumarol FCCP

44
Agentes desacoplantes del gradiente de H+, Ionóforos
Los agentes desacoplantes son sustancias que introducen H+ desde el espacio intermembranoso hacia el interior mitocondrial y disminuyen la fuerza protón-motriz; por lo tanto disminuye la síntesis de ATP.

45
DESACOPLANTE El 2.4-DNF entra en las células en
estado molecular, en el espacio intermembranoso (pH bajo) no se disocia y pasa a la mitocondria, allí hay un pH superior y se disocia, luego introduce H+. UCP1 TERMOGENINA
 no se. disocia y pasa a la mitocondria, allí. hay un pH superior y se disocia, luego introduce H+. UCP1 TERMOGENINA.")
46
Humana cosa es tener compasión de los afligidos; y esto, que en toda persona parece bien, debe máximamente exigirse a quienes hubieron menester consuelo y lo encontraron en los demás.

Presentaciones similares










.>")









