Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Respiración aerobia de la glucosa Catabolismo de lípidos
Metabolismo parte II Respiración aerobia de la glucosa Catabolismo de lípidos

2
Respiración aerobia de la glucosa
La respiración es el proceso catabólico en el que se oxida totalmente la glucosa. El aceptor final de electrones es una molécula inorgánica, que en el caso de la respiración aerobia es el oxígeno. La respiración aerobia consta de varias etapas sucesivas: Glucolisis: Descarboxilación oxidativa del piruvato Ciclo de krebs Cadena de transporte electrónico.

3
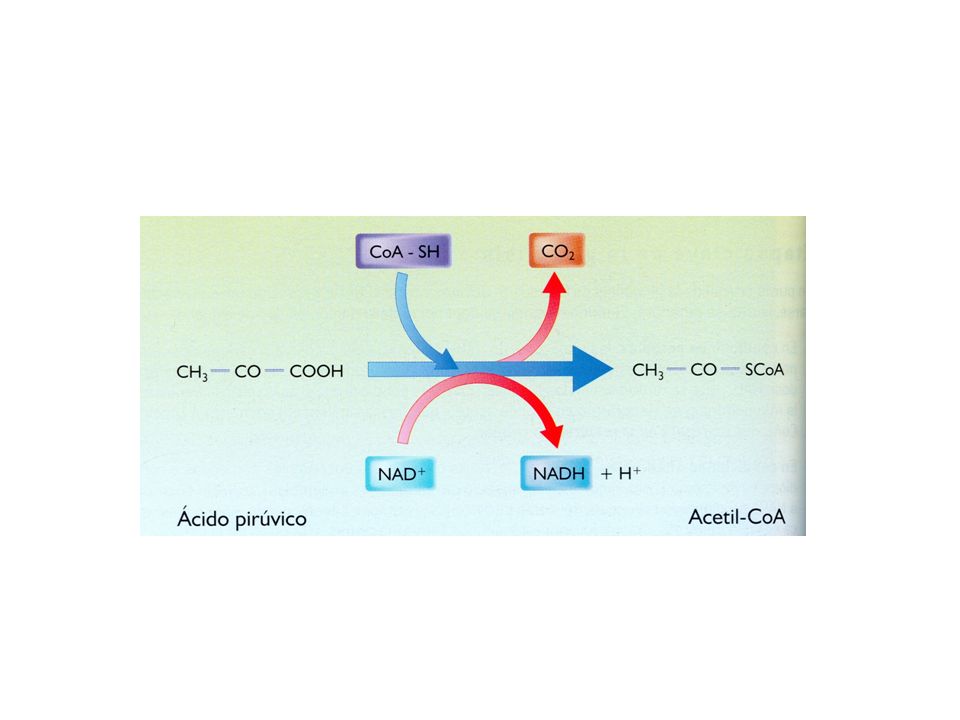
Descarboxilación oxidativa del ácido pirúvico
Compartimento: matriz mitocondrial. El ácido pirúvico procedente de la glucolisis pasa del citosol hacia la matriz mitocondrial. Se produce una descarboxilación oxidativa: Pierde una molécula de CO2 Se oxida formando ácido acético El ácido acético es transferido a una molécula de coenzima A, formando acetil Co A. Estas reacciones están catalizadas por el complejo enzimático piruvato deshidrogenasa. Los electrones liberados son recogidos por el NAD+, que se reduce a NADH

5
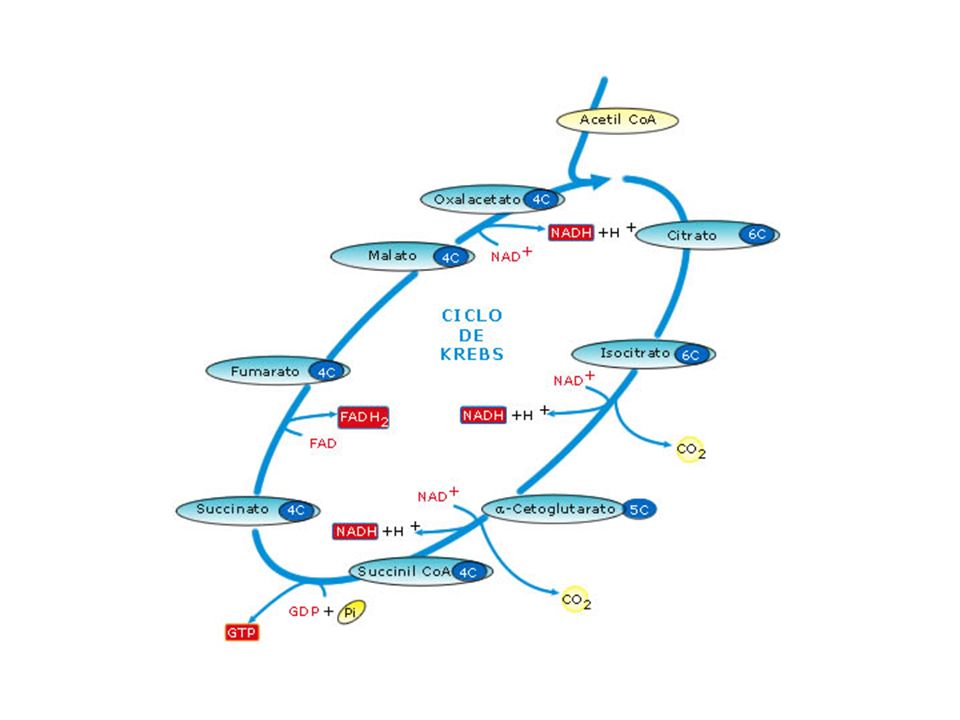
Ciclo de Krebs Es una serie de reacciones que forman una ruta metabólica cíclica. Este ciclo es la ruta final de la oxidación del piruvato, ácidos grasos y cadenas de carbono de los aminoácidos. 2 átomos de carbono del acetil CoA se oxidan totalmente formando 2 moléculas de CO2 Se lleva a cabo en la matriz mitocondrial . Cada reacción es catalizada por una enzima específica.

6
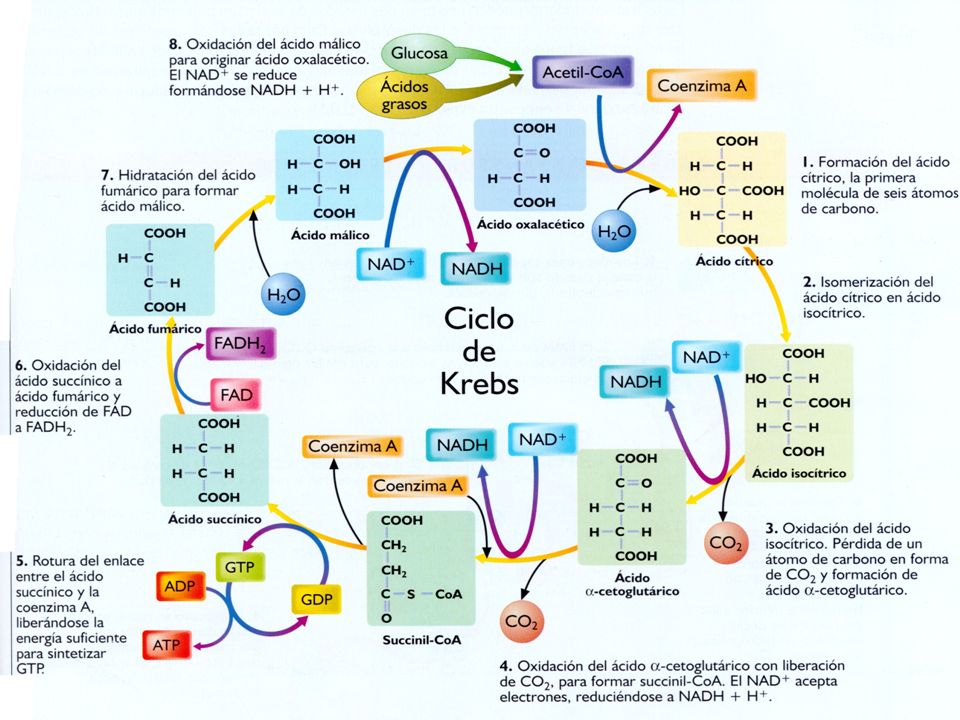
Reacciones del ciclo de krebs
Unión del acetil-S-CoA (2C) con el ácido oxalacético (4C) para formar el ácido cítrico (6C). CITRATO SINTASA. El ácido cítrico se isomeriza a ácido isocítrico. ACONITASA El ácido isocítrico se descarboxila y se oxida perdiendo hidrógenos, con lo que se forma el ácido α-cetoglutárico (5 C). ISOCIRATO DESHIDROGENASA El ácido α-cetoglutárico se descarboxila y deshidrogena, formándose succinil-CoA (4 C) y necesitándose para la reacción la ayuda del CoA. α-CETOGLUTARATO DESHIDROGENSA El succinil-CoA pierde el CoA y se transforma en ácido succínico, liberándose una energía que es suficiente para fosforilar una molécula de GDP y formar una de GTP. SUCCINATO TIOQUINASA El ácido succínico se oxida a ácido fumárico. SUCCINATO DESHIROGENASA El ácido fumárico se hidrata y se transforma en ácido málico. FUMARASA El ácido málico se oxida y se transforma en ácido oxalacético, con lo que se cierra el ciclo. MALATO DESHIDROGENASA
 con el ácido oxalacético (4C) para formar el ácido cítrico (6C). CITRATO SINTASA. El ácido cítrico se isomeriza a ácido isocítrico. ACONITASA. El ácido isocítrico se descarboxila y se oxida perdiendo hidrógenos, con lo que se forma el ácido α-cetoglutárico (5 C). ISOCIRATO DESHIDROGENASA. El ácido α-cetoglutárico se descarboxila y deshidrogena, formándose succinil-CoA (4 C) y necesitándose para la reacción la ayuda del CoA. α-CETOGLUTARATO DESHIDROGENSA. El succinil-CoA pierde el CoA y se transforma en ácido succínico, liberándose una energía que es suficiente para fosforilar una molécula de GDP y formar una de GTP. SUCCINATO TIOQUINASA. El ácido succínico se oxida a ácido fumárico. SUCCINATO DESHIROGENASA. El ácido fumárico se hidrata y se transforma en ácido málico. FUMARASA. El ácido málico se oxida y se transforma en ácido oxalacético, con lo que se cierra el ciclo. MALATO DESHIDROGENASA.")
9
Como en el ciclo de Krebs penetra un compuesto de dos C (el acetil-S-CoA) y se producen dos descarboxilaciones (pasos 3 y 4), la molécula queda totalmente degradada. Además, como en la glucólisis se forman dos moléculas de ácido pirúvico, para la degradación total de una molécula de glucosa son necesarias dos vueltas del ciclo de Krebs. Los GTP se transforman posteriormente en ATP.

10
Balance energético del ciclo de Krebs
Acido Pirúvico Acetil CoA 3 NADH 1 FADH2 1 GTP Como se obtienen 2 piruvatos por cada molécula de glucosa: 2 Acidos Pirúvicos 2 Acetil CoA 6 NADH 2 FADH2 2 GTP

11
Cadena de transporte de electrones. Cadena respiratoria
Es un conjunto de complejos proteicos situados en la membrana mitocondrial interna. Realizan el transporte de electrones desde los coenzimas reducidos (NADH, FADH2) hasta el oxígeno (respiración aerobia, el aceptor final de electrones es el O2, que se reduce formando H2O) En la cadena los electrones fluyen a favor de potencial de oxidorreducción.
 hasta el oxígeno (respiración aerobia, el aceptor final de electrones es el O2, que se reduce formando H2O) En la cadena los electrones fluyen a favor de potencial de oxidorreducción.")
12
Complejos proteicos de la cadena respiratoria
Complejo I: NADH Deshidrogenasa Coenzima Q. Complejo II: Succinato deshidrogenasa Complejo III: Citocromo b-c1 Complejo IV: citocromo oxidasa

13
La energía que se libera cuando pasan los electrones se utiliza para bombear protones desde la matriz mitocondrial al espacio intermembrana.

14
Complejo I: NADH deshidrogenasa
Complejo I: NADH deshidrogenasa. Acepta los electrones del NADH y los pasa al coenzima Q El coenzima Q recibe los electrones de el complejo I y del complejo II. Complejo II, recibe los electrones del FADH2 y los cede al coenzima Q El coenzima Q pasa los electrones al complejo III, después al citocromo c y al complejo IV. Desde el complejo IV pasan los electrones al O2, que junto con los H+ de la matriz se forma H2O.

15
Los coenzimas reducidos se van a oxidar en la última etapa de la respiración, la cadena respiratoria. Pasan por una cadena en la que se reducen y se oxidan diferentes moléculas, a medida que se van traspasando unas a otras los protones y los electrones procedentes del NADH y del FADH2. FADH2

16
Cada transportador tiene un potencial de reducción (tendencia a dar electrones) inferior al anterior (y por tanto se oxida al ceder los e-) y superior al siguiente (se reduce al aceptar los e-). Como la energía liberada durante la oxidación es mayor que la consumida para la reducción, en cada paso hay un sobrante de energía que se invierte en la síntesis de ATP. .

17
Se crea un gradiente electroquímico.
Fosforilación oxidativa. Hipótesis quimiosmótica de Mitchell. Según esta teoría, la energía liberada se invierte en provocar un bombeo de protones (H+) desde la matriz mitocondrial al espacio intermembrana. Se crea un gradiente electroquímico. Cuando los protones (H+) en exceso en el espacio intermembranoso vuelven a la matriz mitocondrial, lo hacen atravesando las partículas F o complejos enzimáticos ATP-sintasa, suministrándoles la energía necesaria para la síntesis de ATP. A partir de un NADH+H+ que ingresa en la cadena respiratoria se obtienen 3ATP. A partir de un FADH2 sólo se obtienen 2ATP, ya que el FADH2 se incorpora a la cadena respiratoria en el complejo coenzima Q reductasa. Al final de la cadena respiratoria aeróbica los hidrógenos se unen al oxígeno y forman agua.
 desde la matriz mitocondrial al espacio intermembrana. Se crea un gradiente electroquímico. Cuando los protones (H+) en exceso en el espacio intermembranoso vuelven a la matriz mitocondrial, lo hacen atravesando las partículas F o complejos enzimáticos ATP-sintasa, suministrándoles la energía necesaria para la síntesis de ATP. A partir de un NADH+H+ que ingresa en la cadena respiratoria se obtienen 3ATP. A partir de un FADH2 sólo se obtienen 2ATP, ya que el FADH2 se incorpora a la cadena respiratoria en el complejo coenzima Q reductasa. Al final de la cadena respiratoria aeróbica los hidrógenos se unen al oxígeno y forman agua.")
19
Partículas F Forman canales por donde pasan los protones
Están formados por: Complejo enzimático F1 Zona de anclaje Fo Por cada 3 protones que pasan se forma una molécula de ATP

20
Rendimiento energético del catabolismo por respiración de la glucosa:
En la glucólisis por cada molécula de glucosa que es degradada se forman 2 moléculas de ácido pirúvico, 2 NADH y 2 ATP. En el sistema piruvato-deshidrogenasa y en el ciclo de Krebs se producen 1 GTP (equivalente a 1 ATP), 4 NADH y 1 FADH2. Si las dos moléculas de ácido pirúvico obtenidas en la glucólisis inician el ciclo de Krebs, todos los productos de éste hay que multiplicarlos por dos: 2 GTP (equivalente a 2 ATP), 8 NADH y 2 FADH2. Las coenzimas reducidas ingresan en la cadena respiratoria y se forma ATP.
, 4 NADH y 1 FADH2. Si las dos moléculas de ácido pirúvico obtenidas en la glucólisis inician el ciclo de Krebs, todos los productos de éste hay que multiplicarlos por dos: 2 GTP (equivalente a 2 ATP), 8 NADH y 2 FADH2. Las coenzimas reducidas ingresan en la cadena respiratoria y se forma ATP.")
21
Descarboxilación oxidativa 2 PIRUVATO 2 ACETIL CoA 2 NADH
GLUCOLISIS PIRUVATO 2 NADH 2 ATP GLUCOSA Descarboxilación oxidativa 2 PIRUVATO 2 ACETIL CoA 2 NADH Ciclo de krebs 2 acetil Co A 6 NADH 2 GTP 2 FADH2 GLUCOLISIS 38 ATP 8 ATP Descarboxilación oxidativa GLUCOSA 6 ATP Ciclo de krebs 24 ATP

22
Transporte electrónico
Rendimiento energético del catabolismo por respiración de la glucosa: Proceso Citoplasma Matriz mitocondrial Transporte electrónico Total Glucolisis 2 ATP 2 NADH 2x (3 ATP) 6 ATP Respiración Ac. Piruvico a Ac. CoA 2 X (1NADH) 2 x (3ATP) Ciclo de Krebs 2 X (1 GTP) 2 X(3 NADH) 2 X(1 FADH2) 6 x (3ATP) 2 x (2ATP) 18 ATP 4 ATP Balance energético global (por molécula de glucosa) 38 ATP
 6 ATP. Respiración. Ac. Piruvico a Ac. CoA. 2 X (1NADH) 2 x (3ATP) Ciclo de Krebs. 2 X (1 GTP) 2 X(3 NADH) 2 X(1 FADH2) 6 x (3ATP) 2 x (2ATP) 18 ATP. 4 ATP. Balance energético global (por molécula de glucosa) 38 ATP.")
23
CATABOLISMO DE OTROS NUTRIENTES
Muchos organismos dependen de otros nutrientes, además de la glucosa. Los seres humanos y otros animales obtienen más energía mediante la oxidación de los ácidos grasos provenientes de los triglicéridos que de la oxidación de la glucosa. En algunos casos también los aminoácidos de las proteínas pueden ser utilizados como combustible. Estos nutrientes se transforman en alguno de los intermediarios que intervienen en la glucólisis o en el ciclo del ácido cítrico.

24
Catabolismo de los lípidos
Una de las funciones de los lípidos es energética (triacilglicéridos). Los acilglicéridos actúan como reserva energética (tanto en animales como en vegetales). En animales los triacilglicéridos se acumulan en el tejido adiposo. Los triacilglicéridos son hidrolizados en el citosol por enzimas llamadas lipasas, que separan los ácidos grasos de la glicerina.
. Los acilglicéridos actúan como reserva energética (tanto en animales como en vegetales). En animales los triacilglicéridos se acumulan en el tejido adiposo. Los triacilglicéridos son hidrolizados en el citosol por enzimas llamadas lipasas, que separan los ácidos grasos de la glicerina.")
25
Grasas Glicerol Glucólisis Ácidos grasos -oxidación Lipasas

26
El glicerol (glicerina) es fosforilado y luego oxidado a gliceraldehído 3-fosfato, que se incorpora a la glucolisis. Los ácidos grasos deben pasar desde el citosol a la matriz mitocondrial (que es donde se produce la b-oxidación.

27
Transporte de los ácidos grasos del citosol a la matriz mitocondrial
El acido graso ha de pasar a la matriz mitocondrial. Para atravesar las membranas mitocondriales se unen a una proteína transportadora específica llamada carnitina. La carnitina vuelve al citoplasma para reutilizarse.

28
Activación de los ácidos grasos
El ácido graso se une al coenzima A, formando acil graso CoA. Esta reacción consume 2 ATP

29
-oxidación de los ácidos grasos
Tiene lugar en la matriz mitocondrial. El Acil graso CoA empieza el proceso de -oxidación

30
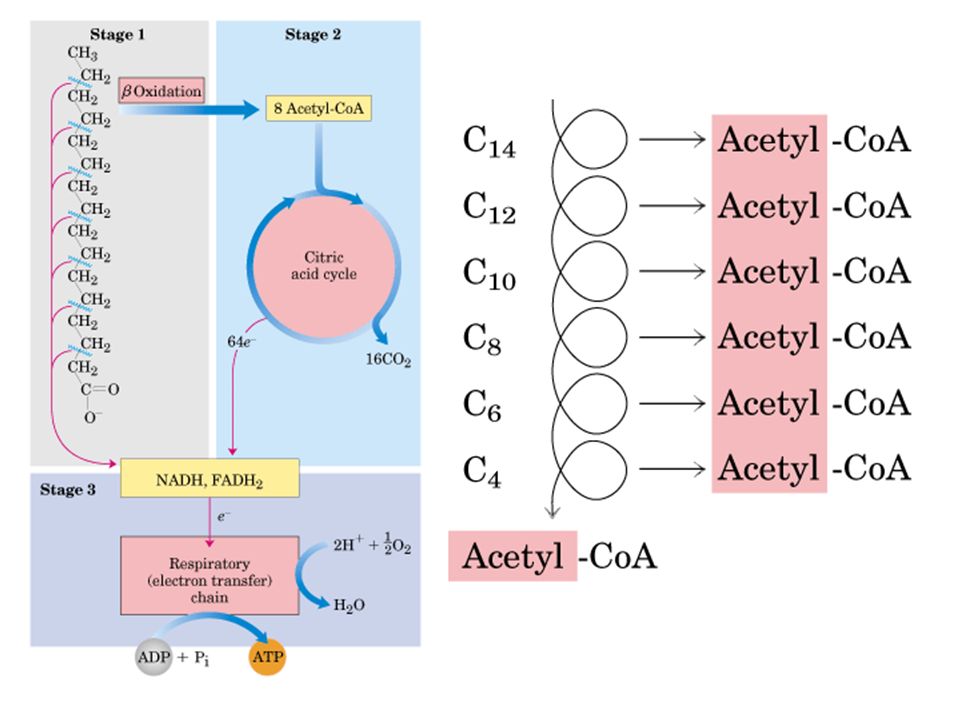
CH3-CH2-(CH2)n-CH2-CH2-COOH
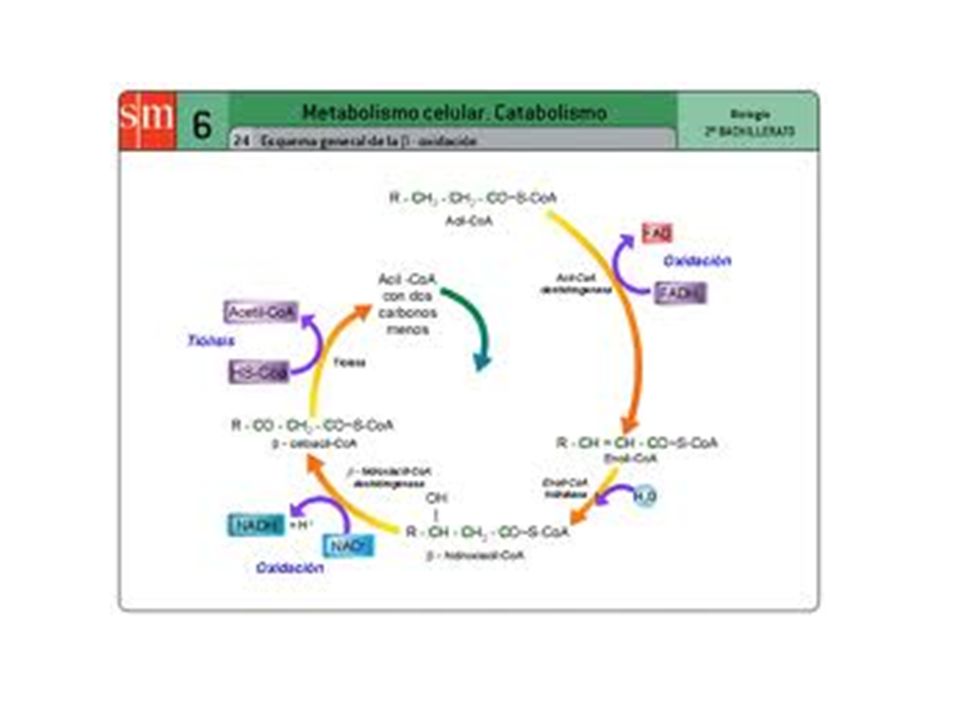
Los ácidos grasos son convertidos en unidades de acetil Co A en un proceso que se denomina -oxidación porque el carbono destinado a ser oxidado es el carbono (el carbono siguiente al , que es el vecino al grupo ácido). CH3-CH2-(CH2)n-CH2-CH2-COOH El proceso, en cuatro pasos da como resultado un acil CoA con 2 carbonos menos, un acetil CoA que se incorpora al ciclo de Krebs, 1 molécula de FADH2 y 1 molécula de NADH (estas dos últimas pasan a la cadena de transporte electrónico). El Acil CoA comienza un nuevo ciclo y así continúa hasta la total degradación del ácido graso. En general los ácidos grasos de los triglicéridos tienen número par de átomos de carbono, por lo que el número de moléculas de coenzima A producidas es igual a la mitad del número de átomos de carbono del ácido graso sometido a -oxidación.
. CH3-CH2-(CH2)n-CH2-CH2-COOH. El proceso, en cuatro pasos da como resultado un acil CoA con 2 carbonos menos, un acetil CoA que se incorpora al ciclo de Krebs, 1 molécula de FADH2 y 1 molécula de NADH (estas dos últimas pasan a la cadena de transporte electrónico). El Acil CoA comienza un nuevo ciclo y así continúa hasta la total degradación del ácido graso. En general los ácidos grasos de los triglicéridos tienen número par de átomos de carbono, por lo que el número de moléculas de coenzima A producidas es igual a la mitad del número de átomos de carbono del ácido graso sometido a -oxidación.")
31
La b oxidación es un ciclo de reacciones en las que la cadena carbonada de los ácidos grasos experimenta una serie de reacciones sucesivas en las que se van escindiendo unidades de 2 carbonos (acetil CoA). Este acetil CoA se dirige al ciclo de krebs para oxidarse totalmente. Cada vuelta de la b-oxidación produce un acil graso CoA (con 2 átomos de carbono menos), 1 FADH2 y 1NADH (que van a la cadena de transporte)
, 1 FADH2 y 1NADH (que van a la cadena de transporte)")
32
Etapas de la b-oxidación
1. deshidrogenación: es una oxidación por deshidrogenación de los átomos de carbono a y b del acil graso CoA, formando un doble enlace y dando lugar a a-b enoil CoA. Se reduce el FAD, dando FADH2. 2. hidratación: entra una molécula de H2O, produciendo saturación del doble enlace, formando un grupo hidroxilo (OH). Se forma b-hidroxiacil CoA 3. oxidación: la deshidrogenación del carbono b produce b-cetoacil-CoA. Se reduce el NAD+ produciendo NADH. 4. Tiolisis: entra una molécula de Coenzima A y produce la ruptura del enlace entre el carbono a y b del b-cetoacil-CoA. se libera 1 molécula de acetil CoA (entra en el ciclo de krebs) y se produce 1 molécula de acil graso Co A con dos carbonos menos que vuelve a dar otra vuelta en la b-oxidación
. Se forma b-hidroxiacil CoA 3. oxidación: la deshidrogenación del carbono b produce b-cetoacil-CoA. Se reduce el NAD+ produciendo NADH. 4. Tiolisis: entra una molécula de Coenzima A y produce la ruptura del enlace entre el carbono a y b del b-cetoacil-CoA. se libera 1 molécula de acetil CoA (entra en el ciclo de krebs) y se produce 1 molécula de acil graso Co A con dos carbonos menos que vuelve a dar otra vuelta en la b-oxidación")
36
Rendimiento de la oxidación de ácido palmítico (16C) TOTAL
Activación del acido graso Ciclo de Krebs Cadena respiratoria - 2 ATP -2 ATP 8 Acetil Co A 8* ( 3 NADH + 1 FADH2 + 1 GTP) = (12 ATP) * 8 96 ATP 7 NADH 3 ATP * 7 21 ATP 7 FADH2 2 ATP * 7 14 ATP TOTAL 129 ATP
 = (12 ATP) * ATP. 7 NADH. 3 ATP * ATP. 7 FADH2. 2 ATP * ATP. TOTAL. 129 ATP.")
Presentaciones similares