Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Sistemas de Expresión Génica

2
Objetivo Conocer el diseño de una estrategia para expresión de proteínas Conocer los sistemas de expresión génica en bacterias, levaduras e insectos.

3
Diseño de una estrategia para expresión de proteínas
Elección del vector Aplicación u objetivos experimentales con la proteína expresada Información conocida de la proteína. Selección de la estrategia de clonado: solubilidad/localización subcelular/fusión a tags/regulación de la expresión. Preparación del vector corte con ER/desfosforilación/purificación Preparación del inserto de DNA plásmido y/o PCR/corte con ER/purificación Clonado del inserto dentro del vector Ligación/transformación en huesped/identificación. De clones positivos/verificación del producto clonado por secuenciación. Transformación dentro del huesped para expresión. Inducción y optimización de la expresión de la proteina de interés análisis cuantitativos de niveles de expresión y actividad biológica. Scale-up Purificaión de la proteína y análisis funcional

4
Vel de crecimiento/ Costo Modificaciones post-traduccionales
Sistemas de expresión de proteínas Sistema Vel de crecimiento/ Costo Nivel de expresión Modificaciones post-traduccionales E. coli 30 min. Bajo Alto NO Levaduras 90 min Medio glicosilación Alta manosa Células insectos 18-24 hs. glicosilación (largo, contenido manosa) Células mamíferos 24 hs Si
 Células mamíferos. 24 hs. Si.")
5
Sistema de expresión en bacterias

6
Manipulación de la expresión génica en bacterias
El objetivo principal de la ingeniería genética con fines industriales es lograr la correcta expresión a altos niveles del gen clonado en el microorganismo huésped elegido. Sin embargo, la clonación directa (sin manipulación adicional) de un gen en un vector normal (ej. como el pBR322) no suele asegurar que nuestro gen de interés se vaya a expresar bien, sobre todo cuando procede de especies alejadas filogenéticamente. Por esta razón hay que proceder al diseño de vectores especializados y a menudo a modificaciones del gen de interés, con objeto de que este pueda transcribirse y traducirse bien en el microorganismo huésped.
 de un gen en un vector normal (ej. como el pBR322) no suele asegurar que nuestro gen de interés se vaya a expresar bien, sobre todo cuando procede de especies alejadas filogenéticamente. Por esta razón hay que proceder al diseño de vectores especializados y a menudo a modificaciones del gen de interés, con objeto de que este pueda transcribirse y traducirse bien en el microorganismo huésped.")
7
Elementos a considerar para determinar el sistema de expresión adecuado
Naturaleza de las secuencias de inicio y final de la transcripción (promotores-operadores, terminadores) Señales para el comienzo de la traducción Eficiencia de la traducción Estabilidad de la proteína en la célula huésped Localización celular de la proteína a expresar Número de copias del gen clonado. Naturaleza de las secuencias de inicio y final de la transcripción (promotores-operadores, terminadores) Señales para el comienzo de la traducción, especialmente sitios de unión del ribosoma y codón de inicio de la traducción Eficiencia de la traducción, lo que puede obligar a utilizar codones sinónimos adaptados a la abundancia de los correspondientes ARNt del huésped Estabilidad de la proteína en la célula huésped Localización celular de la proteína a expresar: en muchos casos interesa que la proteína se secrete, por lo que el diseño genético debe incluir señales de procesamiento postraduccional adecuadas. Otras veces interesa que la proteína se quede en el espacio periplásmico de la bacteria Gram-negativa huésped, o que se retenga en el citoplasma Número de copias del gen clonado. Cuando se pretende que el gen se exprese a alto nivel, se suele escoger un plásmido de control relajado, es decir, con gran número de copias, mientras que si interesa un nivel bajo de expresión se escoge un plásmido de control estricto (poco número de copias) o se recurre a integrar el gen de interés en el cromosoma
 Señales para el comienzo de la traducción. Eficiencia de la traducción. Estabilidad de la proteína en la célula huésped. Localización celular de la proteína a expresar. Número de copias del gen clonado. Naturaleza de las secuencias de inicio y final de la transcripción (promotores-operadores, terminadores) Señales para el comienzo de la traducción, especialmente sitios de unión del ribosoma y codón de inicio de la traducción. Eficiencia de la traducción, lo que puede obligar a utilizar codones sinónimos adaptados a la abundancia de los correspondientes ARNt del huésped. Estabilidad de la proteína en la célula huésped. Localización celular de la proteína a expresar: en muchos casos interesa que la proteína se secrete, por lo que el diseño genético debe incluir señales de procesamiento postraduccional adecuadas. Otras veces interesa que la proteína se quede en el espacio periplásmico de la bacteria Gram-negativa huésped, o que se retenga en el citoplasma. Número de copias del gen clonado. Cuando se pretende que el gen se exprese a alto nivel, se suele escoger un plásmido de control relajado, es decir, con gran número de copias, mientras que si interesa un nivel bajo de expresión se escoge un plásmido de control estricto (poco número de copias) o se recurre a integrar el gen de interés en el cromosoma.")
8
Lo que debe tener un vector de expresión en E. coli
Un promotor eficiente, dotado de las regiones conservadas –35 (TTGACA o parecida) y –10 (TATAAT o parecida) A corta distancia del promotor, una región que al transcribirse suministre el sitio de entrada al ribosoma: la denominada secuencia de Shine-Dalgarno, que es complementaria del extremo 3’ del ARNr 16S A unos 3-11 pb aguas abajo (dowstream) de la secuencia de Shine-Dalgarno, debería situarse el codón ATG que señala el comienzo de la traducción del ARNm. Este codón lo puede suministrar el gen a clonar, o bien el vector puede disponer de ese codón, seguido de alguna diana que permita la inserción del gen a expresar Finalmente, y situado al lado 3’ de todo lo anterior, una secuencia que funcione como terminador de la transcripción (que en su forma de ARN constituye la típica horquilla por emparejamiento, donde la ARN polimerasa se desliga del ARN recién formado).
 y –10 (TATAAT o parecida) A corta distancia del promotor, una región que al transcribirse suministre el sitio de entrada al ribosoma: la denominada secuencia de Shine-Dalgarno, que es complementaria del extremo 3’ del ARNr 16S. A unos 3-11 pb aguas abajo (dowstream) de la secuencia de Shine-Dalgarno, debería situarse el codón ATG que señala el comienzo de la traducción del ARNm. Este codón lo puede suministrar el gen a clonar, o bien el vector puede disponer de ese codón, seguido de alguna diana que permita la inserción del gen a expresar. Finalmente, y situado al lado 3’ de todo lo anterior, una secuencia que funcione como terminador de la transcripción (que en su forma de ARN constituye la típica horquilla por emparejamiento, donde la ARN polimerasa se desliga del ARN recién formado).")
9
Ejemplos de promotores regulables en E. coli
Derivados del promotor lac Derivados del promotor trp Promotor híbrido artificial tac Promotores derivados del fago Promotor del fago T7 Los promotores regulables de E. coli que se emplean en los vectores de expresión más habituales son promotores fuertes pero que bajo ciertas condiciones están reprimidos por las correspondientes proteínas represoras, lo que suministra la base para inducir o desreprimir el gen adyacente a voluntad. Derivados del promotor lac El promotor del operón de la lactosa (lac) ha servido para diseñar elementos de control de expresión en numerosos vectores. Mientras en el medio no exista lactosa (o un inductor apropiado), el promotor está reprimido por el represor (producto del gen lacI). En el laboratorio, la inducción se suele lograr añadiendo al medio un inductor gratuito (que a diferencia de la lactosa, no se metaboliza, pero se une al represor, inactivándolo). Ejemplo de inductor gratuito es el IPTG (isopropil--D-tiogalactósido). Para que el promotor lac se exprese a alto nivel, tenemos que evitar la represión catabólica: en ausencia de glucosa la proteína CAP se une con AMPc, y el complejo se une cerca del promotor, ayudando a la ARN polimerasa a comenzar la transcripción. En los vectores de expresión se suele emplear un promotor lac mutante, conocido como lacUV5 (con una sustitución en la región –10), que es más potente que el promotor silvestre. Por supuesto, esta variante es reprimible por LacI e inducible por IPTG. Derivados del promotor trp El operón trp contiene los genes estructurales que determinan las enzimas para la síntesis del triptófano. Su promotor se ve reprimido, en presencia del triptófano en el medio, por el complejo represor-triptófano. Si no hay triptófano disponible, el represor queda inactivo, y por lo tanto el operón trp se transcribe. En los vectores basados en el promotor trp, la expresión se logra en medio sin triptófano, mientras que se reprime en presencia del aminoácido, o añadiendo el compuesto ácido 3-indol-acrílico. Promotor híbrido artificial tac En los años 80 se diseñó un “promotor artificial” combinando la región –35 del promotor trp y la –10 del promotor lac, poniéndole de nombre promotor tac. Dicho promotor resultó ser 5 veces más potente que el de trp y 10 veces más que el de lac. A semejanza del promotor de lac, el tac es inducible por IPTG y reprimible por glucosa. Se ha usado mucho en vectores de expresión de alto rendimiento. Se llegan a alcanzar niveles de proteína recombinante del 10-30% de la total. Promotores derivados del fago Se puede usar un sistema de expresión regulable consistente en el promotor PL del fago y su represor cI. En la práctica se recurre al represor mutante cI857, que es un mutante termosensible (ts). De esta manera, podemos cultivar la bacteria recombinante a una temperatura moderada (30ºC) a la que el represor impide la transcripción del gen de interés. Cuando el cultivo alcanza la densidad deseada, subimos la temperatura hasta 42ºC, lo que inactiva al represor, y se logra la expresión a alto nivel de nuestro gen desde el promotor PL Promotor del fago T7 El promotor del fago T7 necesita la ARN polimerasa específica del mismo fago, por lo que si queremos expresar genes bajo ese promotor, hay que clonar simultáneamente el gen de la polimerasa de T7. Normalmente este gen de la polimerasa se inserta en el cromosoma de E. coli o en el profago (la versión insertada del cromosoma del en las células lisogénicas) y bajo el control del promotor de lac. Entonces se procede a transformar las células con la construcción genética dotada del gen de interés aguas abajo del promotor del T7. Cuando añadimos el inductor IPTG, se induce el gen de la polimerasa de T7, proteína que transcribe a gran nivel el gen que hemos clonado.
 ha servido para diseñar elementos de control de expresión en numerosos vectores. Mientras en el medio no exista lactosa (o un inductor apropiado), el promotor está reprimido por el represor (producto del gen lacI). En el laboratorio, la inducción se suele lograr añadiendo al medio un inductor gratuito (que a diferencia de la lactosa, no se metaboliza, pero se une al represor, inactivándolo). Ejemplo de inductor gratuito es el IPTG (isopropil--D-tiogalactósido). Para que el promotor lac se exprese a alto nivel, tenemos que evitar la represión catabólica: en ausencia de glucosa la proteína CAP se une con AMPc, y el complejo se une cerca del promotor, ayudando a la ARN polimerasa a comenzar la transcripción. En los vectores de expresión se suele emplear un promotor lac mutante, conocido como lacUV5 (con una sustitución en la región –10), que es más potente que el promotor silvestre. Por supuesto, esta variante es reprimible por LacI e inducible por IPTG. Derivados del promotor trp. El operón trp contiene los genes estructurales que determinan las enzimas para la síntesis del triptófano. Su promotor se ve reprimido, en presencia del triptófano en el medio, por el complejo represor-triptófano. Si no hay triptófano disponible, el represor queda inactivo, y por lo tanto el operón trp se transcribe. En los vectores basados en el promotor trp, la expresión se logra en medio sin triptófano, mientras que se reprime en presencia del aminoácido, o añadiendo el compuesto ácido 3-indol-acrílico. Promotor híbrido artificial tac. En los años 80 se diseñó un promotor artificial combinando la región –35 del promotor trp y la –10 del promotor lac, poniéndole de nombre promotor tac. Dicho promotor resultó ser 5 veces más potente que el de trp y 10 veces más que el de lac. A semejanza del promotor de lac, el tac es inducible por IPTG y reprimible por glucosa. Se ha usado mucho en vectores de expresión de alto rendimiento. Se llegan a alcanzar niveles de proteína recombinante del 10-30% de la total. Promotores derivados del fago Se puede usar un sistema de expresión regulable consistente en el promotor PL del fago y su represor cI. En la práctica se recurre al represor mutante cI857, que es un mutante termosensible (ts). De esta manera, podemos cultivar la bacteria recombinante a una temperatura moderada (30ºC) a la que el represor impide la transcripción del gen de interés. Cuando el cultivo alcanza la densidad deseada, subimos la temperatura hasta 42ºC, lo que inactiva al represor, y se logra la expresión a alto nivel de nuestro gen desde el promotor PL. Promotor del fago T7. El promotor del fago T7 necesita la ARN polimerasa específica del mismo fago, por lo que si queremos expresar genes bajo ese promotor, hay que clonar simultáneamente el gen de la polimerasa de T7. Normalmente este gen de la polimerasa se inserta en el cromosoma de E. coli o en el profago (la versión insertada del cromosoma del en las células lisogénicas) y bajo el control del promotor de lac. Entonces se procede a transformar las células con la construcción genética dotada del gen de interés aguas abajo del promotor del T7. Cuando añadimos el inductor IPTG, se induce el gen de la polimerasa de T7, proteína que transcribe a gran nivel el gen que hemos clonado.")
10
Promotores fuertes y regulables
Expresar un gen a alto nivel, no siempre deberíamos escoger un promotor fuerte y constitutivo. Sin embargo, la mayor parte de las veces no conviene usar tal tipo de promotores, porque al estar expresando permanentemente el gen de interés a alto nivel, pueden sustraerse recursos para el normal crecimiento de las células. Esto es especialmente delicado si la proteína foránea tiene efectos negativos para el huésped. Por lo tanto, lo normal es emplear promotores regulables, que el investigador puede conectar y desconectar a voluntad. Una estrategia habitual es aquella en la que se cultiva la bacteria recombinante hasta que se logran grandes densidades, momento en que se suministra la señal para que el promotor se active y transcriba el gen insertado. En principio, uno podría pensar que si queremos expresar un gen a alto nivel, deberíamos escoger un promotor fuerte y constitutivo. Sin embargo, la mayor parte de las veces no conviene usar tal tipo de promotores, porque al estar expresando permanentemente el gen de interés a alto nivel, pueden sustraerse recursos para el normal crecimiento de las células. Esto es especialmente delicado si la proteína foránea tiene efectos negativos para el huéped. Por lo tanto, lo normal es emplear promotores regulables, que el investigador puede conectar y desconectar a voluntad. Una estrategia habitual es aquella en la que se cultiva la bacteria recombinante hasta que se logran grandes densidades, momento en que se suministra la señal para que el promotor se active y transcriba el gen insertado.

11
Elección del sistema de expresión en E. coli
Tamaño de la proteína: < 100 aminoácidos: estables como proteínas de fusión. >100 aminoácidos: inestables van a cuerpos de inclusión. Cuerpos de inclusión: agregados insolubles intracelulares. Ruptura células (sonicación)--- solubilización y refolding (Urea 8 M)---Diálisis 2) Cantidad de proteína: - Poca cantidad (screening de mutantes): plásmidos de expresión standard - Grandes cantidades: Explorar varios sistemas-huésped y esquemas de purificación. 3) Proteínas activas: considerar la formación de cuerpos de inclusión.
--- solubilización y refolding (Urea 8 M)---Diálisis. 2) Cantidad de proteína: - Poca cantidad (screening de mutantes): plásmidos de expresión standard. - Grandes cantidades: Explorar varios sistemas-huésped y esquemas de purificación. 3) Proteínas activas: considerar la formación de cuerpos de inclusión.")
12
Proteínas expresadas en E. coli
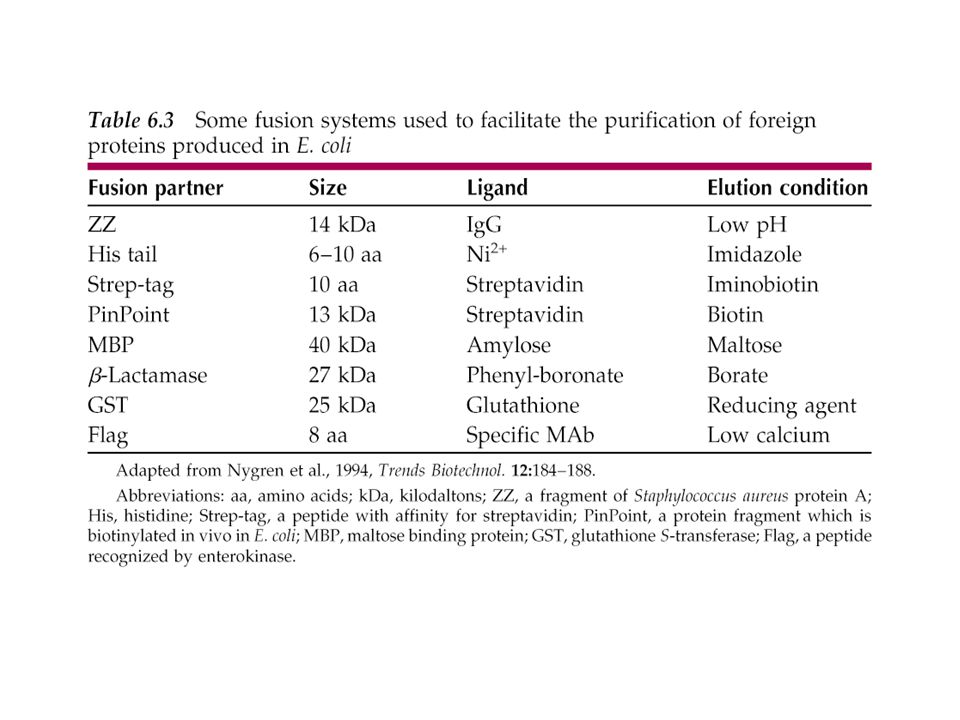
1) Proteínas de fusión: Región que codifica una proteína “tag” fusionada en marco de lectura a la región que codifica para la proteína target. Cuando se transcribe y traduce genera una proteína de fusión purificación protege de proteólisis a la proteína de interés mejora la solubilidad de la proteína estabiliza a la proteína de interés ori Ampr ATG A B MCS Promotor Secuencia “tag”: *Dominio con función enzimática *Señales topogénicas Secuencias consenso para Corte con una proteasa Sitio múltiple de clonado Esquema genérico del vector Proteínas de fusión Cuando uno intenta expresar genes en huéspedes heterólogos es frecuente encontrarse con el hecho de que la proteína buscada se degrade, con lo que se obtienen bajos niveles útiles. Para solventar este problema, uno de los “trucos” consiste en lograr proteínas híbridas, de fusión, en las que nuestra proteína heteróloga (o la parte funcional que nos interesa) está unida covalentemente con una proteína estable del propio huésped, lo que la protege del ataque de proteasas de éste. Para conseguir estas proteínas de fusión, hay que hacer una construcción genética en la que unimos, en la adecuada fase de lectura, un gen del huésped con el gen que queremos expresar. Veamos brevemente los elementos importantes de un vector especializado para obtener proteínas de fusión en E. coli que queden expuestas al exterior desde la membrana externa: Porción terminal 5’ del gen ompF que codifica una proteína de membrana externa. Este segmento suministra las señales de comienzo de la transcripción y de la traducción, así como las señales para que la proteína viaje hasta la membrana externa. Inmediantamente después, una versión truncada del gen lacZ, desprovista no solo del promotor, sino carente de los codones correspondientes a los ocho primeros aminoácidos. A pesar de ello, se podría sintetizar b-galactosidasa funcional en el caso de estar en fase la secuencia que acabamos de describir. Otra característica de este lacZ es que su proteína es capaz de funcionar incluso cuando va fusionada en su extremo amino-terminal con otra proteína. La situación relativa entre el extremo 5’ del ompF y el gen modificado de lacZ en el vector original es tal que este último no está en fase correcta. Por lo tanto, el vector no codifica b-galactosidasa. Ahora bien, si clonamos un gen entre ompF y lacZ de modo que todos queden en fase, se producirá una proteína “tri-híbrida” consistente en el extremo de OmpF fusionado con nuestra proteína, que a su vez va fusionada con la b-galactosidasa. Los clones recombinantes que sinteticen esta proteína tri-híbrida se pueden identificar fácilmente porque tienen actividad b-galactosidasa, detectable como colonias azules en placas provistas de galactósido artificial cromogénico X-gal. La proteína tri-híbrida se puede usar para dos fines: Como antígeno para producir anticuerpos frente a la porción proteica que hemos expresado con el gen clonado Como medio para aislar pequeñas porciones importantes de nuestra proteína. El elemento Flag se compra a una empresa en forma del vector, previsto para su uso en la levadura Saccharomyces cerevisiae. Este elemento determina un péptido de ocho aminoácidos, cuya secuencia es Asp-Tyr-Lys-Asp-Asp-Asp-Asp-Lys. Imaginemos que queremos expresar en levadura el gen de la interleucina 2 (IL-2), una citoquina de gran valor terapéutico. Pues bien, insertamos dicho gen a continuación del elemento genético de Flag. La levadura recombinante produce y secreta la proteína híbrida, que no se degrada. Por otro lado, el péptido Flag nos va a facilitar la tarea de purificar en un solo paso nuestra proteína de interés, por cromatografía de afinidad: Se prepara una columna cromatográfica a base de un soporte sintético (como polipropileno) al que se une anticuerpo monoclonal específico frente al péptido Flag Se hace pasar el extracto o sobrenadante de la cepa recombinante por la columna. La proteína híbrida que nos interesa queda retenida (al engarzarse con el anticuerpo), mientras que las restantes pasan de largo. Para eluir la proteína recombinante, se lava la columna con un tampón bajo en calcio. Aunque la IL-2 recombinante provista de los 8 aminoácidos de Flag en su extremo N-terminal es funcional, las autoridades sanitarias obligan a eliminar ese péptido, cosa que se hace tratando el híbrido con una proteasa específica (enteroquinasa intestinal bovina). Esto último nos ilustra el principio de que para satisfacer los estándares de seguridad y calidad, las proteínas recombinantes logradas como híbridos deben ser despojadas de las porciones del elemento de fusión. Por ello, los elementos de fusión suelen ser secuencias reconocidas por proteasas específicas que cortan en la juntura entre dicho elemento y la proteína de interés, sin atacar a esta.
 Proteínas de fusión: Región que codifica una proteína tag fusionada en marco de lectura a la región que codifica para la proteína target. Cuando se transcribe y traduce genera una proteína de fusión. purificación. protege de proteólisis a la proteína de interés. mejora la solubilidad de la proteína. estabiliza a la proteína de interés. ori. Ampr. ATG. A. B. MCS. Promotor. Secuencia tag : *Dominio con función enzimática. *Señales topogénicas. Secuencias consenso para. Corte con una proteasa. Sitio múltiple de clonado. Esquema genérico del vector. Proteínas de fusión. Cuando uno intenta expresar genes en huéspedes heterólogos es frecuente encontrarse con el hecho de que la proteína buscada se degrade, con lo que se obtienen bajos niveles útiles. Para solventar este problema, uno de los trucos consiste en lograr proteínas híbridas, de fusión, en las que nuestra proteína heteróloga (o la parte funcional que nos interesa) está unida covalentemente con una proteína estable del propio huésped, lo que la protege del ataque de proteasas de éste. Para conseguir estas proteínas de fusión, hay que hacer una construcción genética en la que unimos, en la adecuada fase de lectura, un gen del huésped con el gen que queremos expresar. Veamos brevemente los elementos importantes de un vector especializado para obtener proteínas de fusión en E. coli que queden expuestas al exterior desde la membrana externa: Porción terminal 5’ del gen ompF que codifica una proteína de membrana externa. Este segmento suministra las señales de comienzo de la transcripción y de la traducción, así como las señales para que la proteína viaje hasta la membrana externa. Inmediantamente después, una versión truncada del gen lacZ, desprovista no solo del promotor, sino carente de los codones correspondientes a los ocho primeros aminoácidos. A pesar de ello, se podría sintetizar b-galactosidasa funcional en el caso de estar en fase la secuencia que acabamos de describir. Otra característica de este lacZ es que su proteína es capaz de funcionar incluso cuando va fusionada en su extremo amino-terminal con otra proteína. La situación relativa entre el extremo 5’ del ompF y el gen modificado de lacZ en el vector original es tal que este último no está en fase correcta. Por lo tanto, el vector no codifica b-galactosidasa. Ahora bien, si clonamos un gen entre ompF y lacZ de modo que todos queden en fase, se producirá una proteína tri-híbrida consistente en el extremo de OmpF fusionado con nuestra proteína, que a su vez va fusionada con la b-galactosidasa. Los clones recombinantes que sinteticen esta proteína tri-híbrida se pueden identificar fácilmente porque tienen actividad b-galactosidasa, detectable como colonias azules en placas provistas de galactósido artificial cromogénico X-gal. La proteína tri-híbrida se puede usar para dos fines: Como antígeno para producir anticuerpos frente a la porción proteica que hemos expresado con el gen clonado. Como medio para aislar pequeñas porciones importantes de nuestra proteína. El elemento Flag se compra a una empresa en forma del vector, previsto para su uso en la levadura Saccharomyces cerevisiae. Este elemento determina un péptido de ocho aminoácidos, cuya secuencia es Asp-Tyr-Lys-Asp-Asp-Asp-Asp-Lys. Imaginemos que queremos expresar en levadura el gen de la interleucina 2 (IL-2), una citoquina de gran valor terapéutico. Pues bien, insertamos dicho gen a continuación del elemento genético de Flag. La levadura recombinante produce y secreta la proteína híbrida, que no se degrada. Por otro lado, el péptido Flag nos va a facilitar la tarea de purificar en un solo paso nuestra proteína de interés, por cromatografía de afinidad: Se prepara una columna cromatográfica a base de un soporte sintético (como polipropileno) al que se une anticuerpo monoclonal específico frente al péptido Flag. Se hace pasar el extracto o sobrenadante de la cepa recombinante por la columna. La proteína híbrida que nos interesa queda retenida (al engarzarse con el anticuerpo), mientras que las restantes pasan de largo. Para eluir la proteína recombinante, se lava la columna con un tampón bajo en calcio. Aunque la IL-2 recombinante provista de los 8 aminoácidos de Flag en su extremo N-terminal es funcional, las autoridades sanitarias obligan a eliminar ese péptido, cosa que se hace tratando el híbrido con una proteasa específica (enteroquinasa intestinal bovina). Esto último nos ilustra el principio de que para satisfacer los estándares de seguridad y calidad, las proteínas recombinantes logradas como híbridos deben ser despojadas de las porciones del elemento de fusión. Por ello, los elementos de fusión suelen ser secuencias reconocidas por proteasas específicas que cortan en la juntura entre dicho elemento y la proteína de interés, sin atacar a esta.")
13
Construcción de una proteína de fusión
Fusion partner gene of interest MET ... LEU ARG THR MET VAL ILE ... End ATG ... GTG CGA ACC ATG GTG ATC ... TAG Nco I Note: translation stop of fusion partner gene must be removed Note: reading frame of fusion protein must be contiguous Secuencias consenso para el corte con una proteasa Clivan una secuencia corta y definida de aa La secuencia de clivado para la proteasa está entre el gen carrier y el gen de interés protease cleavage sequence fusion partner gene of interest

14
Esquema general purificación de proteínas de fusión:GST
2- Glutation 1- 3- 4- Elution Corte con proteasa

16
Optimización de la expresión de proteínas en E. coli
Iniciación de la traducción: Promotor fuerte producirá altos niveles de mRNA Se requiere optimizar la traducción. UAAGGAGG AUUCCUCC AUG 5’ 3’ mRNA 16S rRNA small ribosomal subunit Secuencia de Shine Dalgarno: Secuencia complementaria a región en 16S rRNA (subunidad pequeña del ribosoma) Secuencia 6-8 nt a -10 nt del AUG Como se sabe, no todos los ARNm se traducen con la misma eficiencia. Uno de los factores que condicionan esta eficacia diferencial es la presencia, al comienzo del ARNm, de una secuencia (de 6 a 8 bases) que se puede emparejar con otra secuencia complementaria situada en el extremo 3’ del ARNr 16S. Así pues, el ARNm, para que sea traducido bien debe contar con este sitio de unión al ribosoma (RBS en siglas inglesas, también conocido como secuencia de Shine-Dalgarno), que suele consistir en 5’-UAAGGAGG-3’. Por esta razón, muchos vectores de expresión en E. coli se han diseñado para dotar al gen clonado de una secuencia de este tipo, con objeto de que su ARNm se traduzca eficientemente. Aparte de esto, hay que tener presente otras consideraciones: Debe existir una cierta distancia (en torno a 8 bases) entre la secuencia de Shine-Dalgarno y el codón AUG que señala el comienzo de la traducción El segmento inicial del ADN que contiene la secuencia de Shine-Dalgarno y los primeros codones del gen clonado debe ser tal que, una vez transcrito, el ARN no forme emparejamientos intracatenarios. Es decir, a nivel de ARN ese segmento debe de carecer de la capacidad de formar estructuras secundarias en horquillas, que dificultan el acceso de los ribosomas. Otro factor que puede condicionar la eficiencia de traducción deriva del hecho de que nuestro gen de elección tenga un “sesgo de codones” diferente al del organismo huésped, es decir, tenga abundantes codones para los que el anfitrión no disponga de suficiente nivel de los correspondientes ARNt. (Por ejemplo, de los cuatro codones para Gly, el GGA se usa poco en E. coli, pero mucho en humanos; por lo tanto, si queremos expresar un gen humano con abundantes codones GGA en esta bacteria, nos podemos encontrar que la traducción es poco eficiente, debido a la escasez de ARNt que reconozca este triplete). Cuando se nos presenta un problema de este tipo de sesgo de uso de codones, lo único que podemos hacer (aunque suele ser complicado) es sintetizar en laboratorio una versión sinónima de nuestro gen en la que los codones estén “optimizados” al uso del huésped.
 Secuencia 6-8 nt a -10 nt del AUG. Como se sabe, no todos los ARNm se traducen con la misma eficiencia. Uno de los factores que condicionan esta eficacia diferencial es la presencia, al comienzo del ARNm, de una secuencia (de 6 a 8 bases) que se puede emparejar con otra secuencia complementaria situada en el extremo 3’ del ARNr 16S. Así pues, el ARNm, para que sea traducido bien debe contar con este sitio de unión al ribosoma (RBS en siglas inglesas, también conocido como secuencia de Shine-Dalgarno), que suele consistir en 5’-UAAGGAGG-3’. Por esta razón, muchos vectores de expresión en E. coli se han diseñado para dotar al gen clonado de una secuencia de este tipo, con objeto de que su ARNm se traduzca eficientemente. Aparte de esto, hay que tener presente otras consideraciones: Debe existir una cierta distancia (en torno a 8 bases) entre la secuencia de Shine-Dalgarno y el codón AUG que señala el comienzo de la traducción. El segmento inicial del ADN que contiene la secuencia de Shine-Dalgarno y los primeros codones del gen clonado debe ser tal que, una vez transcrito, el ARN no forme emparejamientos intracatenarios. Es decir, a nivel de ARN ese segmento debe de carecer de la capacidad de formar estructuras secundarias en horquillas, que dificultan el acceso de los ribosomas. Otro factor que puede condicionar la eficiencia de traducción deriva del hecho de que nuestro gen de elección tenga un sesgo de codones diferente al del organismo huésped, es decir, tenga abundantes codones para los que el anfitrión no disponga de suficiente nivel de los correspondientes ARNt. (Por ejemplo, de los cuatro codones para Gly, el GGA se usa poco en E. coli, pero mucho en humanos; por lo tanto, si queremos expresar un gen humano con abundantes codones GGA en esta bacteria, nos podemos encontrar que la traducción es poco eficiente, debido a la escasez de ARNt que reconozca este triplete). Cuando se nos presenta un problema de este tipo de sesgo de uso de codones, lo único que podemos hacer (aunque suele ser complicado) es sintetizar en laboratorio una versión sinónima de nuestro gen en la que los codones estén optimizados al uso del huésped.")
17
2) Uso de codones: El código genético es redundante, usa 61 codones para 20 aa.
Los codones (excepto Met y Trp con 1 codón c/u) que codifican para un aa no son usados con igual frecuencia. Algunos codones usados infrecuentemente=↓[RNAt] Codon Aminoácido Frec. uso en E. coli Frec. uso en humanos GAG Glutámico 0,3 0,59 GAA 0,7 0,41 CCG Prolina 0,55 0,11 CCA 0,20 0,27 CCU 0,16 0,29 CCC 0,10 0,33 Cepa RosettaTM: cepa diseñada para aumentar la expresión de proteínas eucariotas que poseen codones de baja frecuencia en E.coli.
 que codifican para un aa no son. usados con igual frecuencia. Algunos codones usados infrecuentemente=↓[RNAt] Codon. Aminoácido. Frec. uso en E. coli. Frec. uso en humanos. GAG. Glutámico. 0,3. 0,59. GAA. 0,7. 0,41. CCG. Prolina. 0,55. 0,11. CCA. 0,20. 0,27. CCU. 0,16. 0,29. CCC. 0,10. 0,33. Cepa RosettaTM: cepa diseñada para aumentar la expresión de proteínas eucariotas que poseen codones de baja frecuencia en E.coli.")
18
3) Estructura del mRNA: Región del codón de iniciación AUG en extremo 5’ del mRNA no debe ser complementaria a sí misma, podría bloquear el movimiento del ribosoma y evitar la traducción 4) Cepas deficientes en proteasas: Lon y OmpT
 Cepas deficientes en proteasas: Lon y OmpT.")
19
Elección del promotor Promotores basados en el operón Lac: 1) Promotor Lac
 Promotor Lac")
20
Lac promoter pGEM®-3Z Vector sequence reference points.
T7 RNA polymerase transcription initiation site 1; multiple cloning region 5-61 SP6 RNA polymerase promoter (.17 to +3) 67-86; SP6 RNA polymerase transcription initiation site 69 lac operon sequences ; binding site of pUC/M13 Reverse Sequencing Primer lacZ start codon 108; lacZ operator β-lactamase (Ampr) coding region binding site of pUC/M13 Forward Sequencing Primer T7 RNA polymerase promoter (-17 to +3) Cepa huesped: lacZDM15. Expresa LacIq
 67-86; SP6 RNA polymerase transcription initiation site 69. lac operon sequences ; binding site of pUC/M13 Reverse Sequencing Primer lacZ start codon 108; lacZ operator β-lactamase (Ampr) coding region binding site of pUC/M13 Forward Sequencing Primer T7 RNA polymerase promoter (-17 to +3) Cepa huesped: lacZDM15. Expresa LacIq.")
21
Híbrido de promotores lac y trp Regulado por el represor lac
2) Promotores tac y trc: 3x más fuertes que trp y 10x más fuertes que lac Híbrido de promotores lac y trp Regulado por el represor lac Independiente de la regulación por cAMP XXXXXXXXXXXXXXXXXX -35 promotor trp -10 promotor lac Separación 16 pb=promotor tac 17 pb=promotor trc Gen de interés
 Promotores tac y trc: 3x más fuertes que trp y 10x más fuertes que lac. Híbrido de promotores lac y trp. Regulado por el represor lac. Independiente de la regulación por cAMP. XXXXXXXXXXXXXXXXXX. -35 promotor trp. -10 promotor lac. Separación. 16 pb=promotor tac. 17 pb=promotor trc. Gen de interés.")
22
Ptac promoter : terminador de la transcripción Promotor Ptac/IPTG:
Polylinker: MalE: secuencia señal de la proteína Maltosa Binding Protein (MBP), periplásmica con alta afinidad por maltosa.
, periplásmica con alta afinidad por maltosa.")
23
El gen de interés puede estar bajo el control del promotor del gen 10
3) Promotores basados en el gen 10 del bacteriófago T7: Todos los promotores del fago T7 (incluído el gen 10) requieren la T7 RNA Polimerasa para expresarse Inducer T7 RNA Pol El gen de la T7 RNA Pol puede estar bajo el control del promotor lac en genómico de E.coli lac pro T7 RNA Pol Protein of Interest T7 RNA Pol El gen de interés puede estar bajo el control del promotor del gen 10 g10 pro gene of interest
 Promotores basados en el gen 10 del bacteriófago T7: Todos los promotores del fago T7 (incluído el gen 10) requieren la T7 RNA Polimerasa para expresarse. Inducer. T7 RNA Pol. El gen de la T7 RNA Pol puede estar bajo el control del promotor lac en genómico de E.coli. lac pro. T7 RNA Pol. Protein of. Interest. T7 RNA Pol. El gen de interés puede estar bajo el control del promotor del gen 10. g10 pro. gene of interest.")
24
T7 promoter

25
pRSET Expression Vector
The pRSET vector is designed for high-level prokaryotic expression controlled by the strong bacteriophage T7 promoter. Expression is induced by the production of T7 RNA polymerase in the BL21(DE3)pLysS host E. coli cells. These cells also produce T7 lysozyme to reduce basal expression of target genes. The pRSETvector offers: High-level expression from the bacteriophage T7 promoter T7 gene 10 sequence to provide protein stability N-terminal polyhistidine (6xHis) tag for rapid purification with ProBond® resin N-terminal Xpress® epitope for protein detection with the Anti-Xpress® Antibody Enterokinase cleavage site for removal of fusion tag f1 origin for ssDNA rescue to allow easy sequencing and mutagenesis
pLysS host E. coli cells. These cells also produce T7 lysozyme to reduce basal expression of target genes. The pRSETvector offers: High-level expression from the bacteriophage T7 promoter. T7 gene 10 sequence to provide protein stability. N-terminal polyhistidine (6xHis) tag for rapid purification with ProBond® resin. N-terminal Xpress® epitope for protein detection with the Anti-Xpress® Antibody. Enterokinase cleavage site for removal of fusion tag. f1 origin for ssDNA rescue to allow easy sequencing and mutagenesis.")
26
4) Promotores basados en el promotor del bacteriófago l pL:
No tryptophan Transcription Ptrp PO lcI repressor lPL promoter- triptofano Bacterial Chromosome cI repressor No transcription PL PO ATG-GENE pLEX vector Tryptophan trp repressor No transcription Ptrp PO lcI repressor Bacterial Chromosome Transcription PL PO ATG-GENE pLEX vector Mechanism of transcriptional regulation with pLEX

27
Problemas cuando se expresan proteínas eucarióticas en cél procariotas:
Inestables. Sin actividad biológica. Contaminantes procarióticos. Soluciones: se han desarrollado sistemas de expresión en eucariotas Esp. importantes para proteínas terapéuticas. Necesidad de que tengan idénticas propiedades bioquímicas, biofísicas y funcionales a la proteína natural. Modificaciones post-traduccionales Formación de uniones disulfuro correctas. Clivaje proteolítico del precursor inactivo. Glicosilación- agregado de residuos de azúcar Alteración de aminoácidos en la proteína: fosforilación, acetilación, agregado de grupos sulfato, agregado de ácidos grasos

28
Producción de proteínas recombinantes en células eucariotas
Problemas que suelen aparecer cuando proteínas eucarióticas se expresan en cél procariotas inestables sin actividad biológica contaminantes procarióticos

29
Sistemas de expresión eucariotas
Para eliminar los problemas de expresión en procariotas, se han desarrollado sistemas de expresión eucariotas Esp. importantes para proteínas terapéuticas Necesidad de que tengan idénticas propiedades a la proteína natural : bioquímicas biofísicas funcionales

30
Sistemas de expresión eucariotas
Cuál es la diferencia? Modificación post-traduccional de la mayoría de las proteínas eucariotas Cél procariotas no pueden realizar estas modificaciones

31
Modificaciones post-traduccionales
Formación de uniones disulfuro correctas Clivaje proteolítico del precursor inactivo Glicosilación- agregado de residuos de azúcar Alteración de aminoácidos en la proteína fosforilación acetilación agregado de grupos sulfato Agregado de ácidos grasos

32
Sistemas de expresión en levaduras

33
Sistemas de expresión en levaduras: ventajas
Organismo unicelular Bien caracterizadas genética y fisiológicamente Promotores fuertes disponibles Plásmido natural (2µm) Eucariotas, ocurren modificaciones post-transduccionales Segregan algunas proteínas No patógeno
 Eucariotas, ocurren modificaciones post-transduccionales. Segregan algunas proteínas. No patógeno.")
34
Vectores de levaduras YIp: integrativos YEp: episomales
- una copia integrada a un cromosoma YEp: episomales - ori 2m (alto número de copias) YCp: centroméricos ARS, CEN (1 o 2 copias por cél) Todos tienen marcador de auxotrofia y son shuttle
 YCp: centroméricos. ARS, CEN (1 o 2 copias por cél) Todos tienen marcador de auxotrofia y son shuttle.")
35
Vectores episomales Son ampliamente usados
A menudo inestables en cultivos a gran escala (>10L) Estrategia - alterar las condiciones de crecimiento para estabilizar el vector episomal usar cepas mutantes que requieren nutrientes específicos. Leucina, Triptofano, Histidina el gen que provee el fenotipo salvaje se encuentra en el plásmido episomal
 Estrategia - alterar las condiciones de crecimiento para estabilizar el vector episomal. usar cepas mutantes que requieren nutrientes específicos. Leucina, Triptofano, Histidina. el gen que provee el fenotipo salvaje se encuentra en el plásmido episomal.")
37
Promotores de levaduras
Promoter Status Acid phosphatase PHO5 Inducible Alcohol dehydrogenase I ADH I Constitutive Alcohol dehydrogenase II ADH II Inducible Galctokinase GAL 1 Inducible Metallothionein Cup 1 Inducible Phosphoglycerate kinase PGK Constitutive Triose Phosphate isomerase TPI Constitutive

38
Sistemas de expresión en levaduras: desventajas
Inestabilidad de plásmidos en gran escala Vectores episomales Superglicosilación de glicoproteínas 100+ residuos manosa vs normal puede alterar la actividad de la proteína Proteínas secretadas quedan atrapadas en periplasma (entre la membrana y la pared celular) se usan señales peptídicas
 se usan señales peptídicas.")
39
Soluciones a los problemas en sistemas de expresión en levaduras
Levaduras modificadas genéticamente para llevar a cabo glicosilación = humanos Science Vol 301, p (29 Aug 2003) Deleción de genes de glicosilación de levaduras Agregado de genes de glicosilación humanos
 Deleción de genes de glicosilación de levaduras. Agregado de genes de glicosilación humanos.")
40
Soluciones a los problemas en sistemas de expresión en levaduras
Usar otro tipo de levaduras Pichia pastoris Hansenula polymorpha No sólo Saccharomyces cerevisiae Usar otros eucariotas Cél de insectos Cultivos de cél de mamíferos

41
Levaduras como sistema de expresión
Rápido crecimiento, bajo costo similar a E coli. Modificaciones postraduccionales escenciales para la función proteica. Problema (difieren de eucariotas superiores): forma de N-glicosilación y O-glicosilación: uso de levaduras con N-gycosilación humanizada. Saccharomyces cerevisiae Económico, bioseguro para aplicaciones humanos. Posee características bioquímicas, genéticas, similiar a eucariotas superiores. Es adaptable a fermentación gran escala. Herramienta para estudios de complementación. otros Pichia pastoris y Hansenula polymorpha.
: forma de N-glicosilación y O-glicosilación: uso de levaduras con N-gycosilación humanizada. Saccharomyces cerevisiae. Económico, bioseguro para aplicaciones humanos. Posee características bioquímicas, genéticas, similiar a eucariotas superiores. Es adaptable a fermentación gran escala. Herramienta para estudios de complementación. otros Pichia pastoris y Hansenula polymorpha.")
42
YEp: episomales: ori 2m (alto número de copias)
Vectores de levaduras Tipos de vectores YIp: integrativos: una copia integrada al cromosoma YEp: episomales: ori 2m (alto número de copias) YCp: centroméricos: ARS, CEN (1 o 2 copias por cél) Promotores Promoter Status Acid phosphatase PHO5 Inducible Alcohol dehydrogenase I ADH I Constitutive Alcohol dehydrogenase II ADH II Inducible Galctokinase GAL 1 Inducible Metallothionein Cup 1 Inducible Phosphoglycerate kinase PGK Constitutive Triose Phosphate isomerase TPI Constitutive
 YCp: centroméricos: ARS, CEN (1 o 2 copias por cél) Promotores. Promoter Status. Acid phosphatase PHO5 Inducible. Alcohol dehydrogenase I ADH I Constitutive. Alcohol dehydrogenase II ADH II Inducible. Galctokinase GAL 1 Inducible. Metallothionein Cup 1 Inducible. Phosphoglycerate kinase PGK Constitutive. Triose Phosphate isomerase TPI Constitutive.")
43
Vectores episomales Vectores centroméricos

45
Expresión de proteínas en Pichia pastoris
Requiere técnicas simples para la manipulación genética (similar a S. cerevisiae) Producción de altos niveles de proteínas (intra y extracelular) Modificaciones post-traduccionales: glicosilación, puente disulfuro, procesamiento proteolítico. Kits de expresión disponibles. Empleo del gen AOX1 Ampr, oriE: mantenimiento en E. coli AOX1p: promotor de alcohol oxidasa. AOX1t: secuencias terminador transcripción. MCS: sitio múltiple clonado. HIS4: marcador biosintético. 3´-AOX1 MCS Diagrama general de un vector P pastoris shuttle
 Producción de altos niveles de proteínas (intra y extracelular) Modificaciones post-traduccionales: glicosilación, puente disulfuro, procesamiento proteolítico. Kits de expresión disponibles. Empleo del gen AOX1. Ampr, oriE: mantenimiento en E. coli. AOX1p: promotor de alcohol oxidasa. AOX1t: secuencias terminador transcripción. MCS: sitio múltiple clonado. HIS4: marcador biosintético. 3´-AOX1. MCS. Diagrama general de un vector P pastoris shuttle.")
46
Integración del vector de P. pastoris
Se integran para maximizar la estabilidad del sistema de expresión. Corte del vector que lleva inserto de interés flanqueado por secuencias del gen AOX1 (para recombinación homóloga requiere regiones terminales mas largas que S cerevisiae.) Transformación con vector linealizado por electroporación o métodos químicos. Cepa huésped: his4 pep4 prb1 (auxotrófica para Histidina y deficiente en proteasas). Inserto Selección en medio his- Cepa resultante aox1=Muts Metanol: crec lento, alta producción prot.
 Transformación con vector linealizado por electroporación o métodos químicos. Cepa huésped: his4 pep4 prb1 (auxotrófica para Histidina y deficiente en proteasas). Inserto. Selección en medio his- Cepa resultante aox1=Muts. Metanol: crec lento, alta producción prot.")
47
Pichia methanolica Expression System
Expresión controlada por promotor AUG1 (alcohol oxidasa): reprimido por glicerol o glucosa Inducido por metanol a-factor: péptido señal para secreción de la proteína expresada. V5 epítope: para detección con anticuerpos anti-V5 6xHis: tag polihistidina purificación níquel-agarosa. ADE2: marcador biosintético.
: reprimido por glicerol o glucosa. Inducido por metanol. a-factor: péptido señal para secreción de la proteína expresada. V5 epítope: para detección con anticuerpos anti-V5. 6xHis: tag polihistidina purificación níquel-agarosa. ADE2: marcador biosintético.")
48
Sistemas de expresión en insectos
Los baculovirus son un tipo de virus que infectan artrópodos, especialmente insectos. Durante su infección, las partículas virásicas se empaquetan en unos grupos llamados cuerpos poliédricos, que son cuerpos de inclusión localizados en el núcleo celular, compuestos mayoritariamente de la proteína poliedrina. La poliedrina se sintetiza en grandes cantidades al final del ciclo infectivo, pudiendo llegar a suponer el 50% del total proteico de la célula. El genoma de los baculovirus consta de una molécula circular de ADN de c.d., de un tamaño de entre 88 y 200 kb. Aunque los poliedros se necesitan para la infección, no se requieren para mantener el cultivo celular infectado. Por lo tanto, el gen de la poliedrina es dispensable y puede hacerse hueco en esa parte del genoma para manipulación genética. Como el genoma es demasiado grande para manupular directamente, se recurre a lo siguiente: El ADN heterólogo se inserta en un vector intermedio llamado de transferencia, sin capacidad infectiva, que es un vector con secuencias de E. coli (para hacer las manipulaciones fácilmente en la bacteria), y que lleva el eficiente promotor de la poliedrina, tras el cual se inserta el gen de interés. Se co-transfectan células de insecto con el vector que lleva el ADN clonado y con ADN virásico no recombinante. Tiene lugar la recombinación in vivo entre el vector de transferencia y secuencias del genoma virásico, con lo que se generan genomas recombinantes que se pueden seleccionar y uar para producir la proteína de interés. Presentan las ventajas de que las proteínas foráneas se expresan a alto nivel, experimentando modificaciones postraduccionales similares a las de mamíferos. El coste de producción es superior al de levaduras, pero menor al de usar células de mamíferos Se ha logrado adaptar líneas celulares de insectos a algunos de los requerimientos habituales de la producción a gran escala, incluyendo la resistencia a la agitación de los fermentadores Hoy día ya se pueden usar cultivos de células de insectos en biorreactores parecidos a los empleados con los microorganismos Se han hecho avances en diseño de medios de cultivo más sencillos y baratos, que no necesitan suero. Todo ello permite el escalado, esencial para la producción de grandes cantidades de proteínas terapéuticas recombinantes Una de las aplicaciones más importantes de los baculovirus derivados de cultivos de células de insectos es su uso como insecticidas biológicos selectivos y seguros, que se emplean sobre todo en lucha biológica e integrada frente a plagas que afectan a bosques
, y que lleva el eficiente promotor de la poliedrina, tras el cual se inserta el gen de interés. Se co-transfectan células de insecto con el vector que lleva el ADN clonado y con ADN virásico no recombinante. Tiene lugar la recombinación in vivo entre el vector de transferencia y secuencias del genoma virásico, con lo que se generan genomas recombinantes que se pueden seleccionar y uar para producir la proteína de interés. Presentan las ventajas de que las proteínas foráneas se expresan a alto nivel, experimentando modificaciones postraduccionales similares a las de mamíferos. El coste de producción es superior al de levaduras, pero menor al de usar células de mamíferos. Se ha logrado adaptar líneas celulares de insectos a algunos de los requerimientos habituales de la producción a gran escala, incluyendo la resistencia a la agitación de los fermentadores. Hoy día ya se pueden usar cultivos de células de insectos en biorreactores parecidos a los empleados con los microorganismos. Se han hecho avances en diseño de medios de cultivo más sencillos y baratos, que no necesitan suero. Todo ello permite el escalado, esencial para la producción de grandes cantidades de proteínas terapéuticas recombinantes. Una de las aplicaciones más importantes de los baculovirus derivados de cultivos de células de insectos es su uso como insecticidas biológicos selectivos y seguros, que se emplean sobre todo en lucha biológica e integrada frente a plagas que afectan a bosques.")
49
Que son los baculovirus
Los baculovirus son un grupo de virus encontrados mayormente en insectos (Baculoviridae). Tienen una estructura de varilla (baculum=bastón), un diámetro de entre 40-50 nm y una longitud de entre nm. Su información genética está almacenada en una molécula de ADN doble cadena, circular, de aproximadamente Kpb.
. Tienen una estructura de varilla. (baculum=bastón), un diámetro de entre nm y una longitud de entre nm. Su información genética está almacenada en. una molécula de ADN doble cadena, circular, de aproximadamente Kpb.")
50
Baculovirus como sistema de expresión

51
Características Ambiente eucariota para la producción de proteínas
Expresión excepcionalmente alta Expresión durante la fase de oclusión Expresión a 27ºC Capacidad de grandes inserciones Expresión eficiente de genes no “spliceados” (cDNAs) Simplicidad de la tecnología
 Simplicidad de la tecnología.")
53
Asignacion

Presentaciones similares