Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Autotrofía y heterotrofía. Macronutrientes y micronutrientes
Autotrofía y heterotrofía. Macronutrientes y micronutrientes. Nutrientes universales y particulares. Fijación del CO2. Fijación de N2. Factores de crecimiento. Captación de nutrientes Nutrición microbiana En este tema veremos las formas básicas de aprovisionamiento de nutrientes por parte de los microorganismos. Insistiremos en conceptos básicos como autotrofía, heterotrofía, mixotrofía, etc. Abordaremos los nutrientes universales, los particulares y los factores de crecimiento.

2
Conceptos básicos (I) Nutrición: captación del medio de las sustancias para crecer (= nutrientes). Los nutrientes se necesitan para Fines energéticos (en quimiotrofos) mantenimiento Fines biosintéticos (anabolismo, reacciones plásticas) La nutrición es el proceso por el que los seres vivos toman del medio donde habitan las sustancias químicas que necesitan para crecer. Dichas sustancias se denominan nutrientes, y se requieren para dos objetivos: · fines energéticos (reacciones de mantenimiento); · fines biosintéticos (reacciones plásticas o anabolismo). Las biosíntesis de nuevos componentes celulares son procesos que requieren energía procedente del medio ambiente. En el capítulo anterior vimos los principales modos de captación y obtención de energía existentes en las bacterias.
 mantenimiento. Fines biosintéticos (anabolismo, reacciones plásticas) La nutrición es el proceso por el que los seres vivos toman del medio donde habitan las sustancias químicas que necesitan para crecer. Dichas sustancias se denominan nutrientes, y se requieren para dos objetivos: · fines energéticos (reacciones de mantenimiento); · fines biosintéticos (reacciones plásticas o anabolismo). Las biosíntesis de nuevos componentes celulares son procesos que requieren energía procedente del medio ambiente. En el capítulo anterior vimos los principales modos de captación y obtención de energía existentes en las bacterias.")
3
Conceptos básicos (II)
Punto de vista de los fines de aprovisionamiento de energía: Litotrofía: solo requieren sustancias inorgánicas sencillas donantes de electrones : SH2, S0, NH3, NO2-, Fe2+ Organotrofía: requieren compuestos orgánicos (hidratos de C, hidrocarburos, lípidos, proteínas, alcoholes, etc) Punto de vista biosintético (fuente de C): autotrofía: fijación del CO2 heterotrofía: fuente orgánica de carbono Otros conceptos: autotrofía estricta: no pueden crecer usando materia orgánica mixotrofía: metabolismo energético litotrofo, pero usan fuente orgánica de C para su metabolismo biosintético Es importante tener claro desde el principio una serie de conceptos y nomenclaturas relacionados con los principales tipos de nutrición bacteriana. Puesto que, como acabamos de ver, la nutrición presenta un aspecto de aprovisionamiento de energía y otro de suministro de materiales para la síntesis celular, podemos hablar de dos “clasificaciones” de tipos de nutrición: 1) Desde el punto de vista de los fines de aprovisionamiento de energía, las bacterias se pueden dividir en: a) litotrofas: son aquellas que sólo requieren sustancias inorgánicas sencillas (SH2 S0, NH3, NO2-, Fe, etc.). b) organotrofas: requieren compuestos orgánicos (hidratos de carbono, hidrocarburos, lípidos, proteínas, alcoholes...). 2) Desde el punto de vista biosintético (o sea, para sus necesidades plásticas o de crecimiento), las bacterias se pueden dividir en: a) autotrofas: crecen sintetizando sus materiales a partir de sustancias inorgánicas sencillas. Ahora bien, habitualmente el concepto de autotrofía se limita a la capacidad de utilizar una fuente inorgánica de carbono, a saber, el CO2. b) heterotrofas: su fuente de carbono es orgánica (si bien otros elementos distintos del C pueden ser captados en forma inorgánica). Otros conceptos: autotrofas estrictas son aquellas bacterias incapaces de crecer usando materia orgánica como fuente de carbono. Mixotrofas son aquellas bacterias con metabolismo energético litotrofo (obtienen energía de compuestos inorgánicos), pero requieren sustancias orgánicas como nutrientes para su metabolismo biosintético.
 Punto de vista biosintético (fuente de C): autotrofía: fijación del CO2. heterotrofía: fuente orgánica de carbono. Otros conceptos: autotrofía estricta: no pueden crecer usando materia orgánica. mixotrofía: metabolismo energético litotrofo, pero usan fuente orgánica de C para su metabolismo biosintético. Es importante tener claro desde el principio una serie de conceptos y nomenclaturas relacionados con los principales tipos de nutrición bacteriana. Puesto que, como acabamos de ver, la nutrición presenta un aspecto de aprovisionamiento de energía y otro de suministro de materiales para la síntesis celular, podemos hablar de dos clasificaciones de tipos de nutrición: 1) Desde el punto de vista de los fines de aprovisionamiento de energía, las bacterias se pueden dividir en: a) litotrofas: son aquellas que sólo requieren sustancias inorgánicas sencillas (SH2 S0, NH3, NO2-, Fe, etc.). b) organotrofas: requieren compuestos orgánicos (hidratos de carbono, hidrocarburos, lípidos, proteínas, alcoholes...). 2) Desde el punto de vista biosintético (o sea, para sus necesidades plásticas o de crecimiento), las bacterias se pueden dividir en: a) autotrofas: crecen sintetizando sus materiales a partir de sustancias inorgánicas sencillas. Ahora bien, habitualmente el concepto de autotrofía se limita a la capacidad de utilizar una fuente inorgánica de carbono, a saber, el CO2. b) heterotrofas: su fuente de carbono es orgánica (si bien otros elementos distintos del C pueden ser captados en forma inorgánica). Otros conceptos: autotrofas estrictas son aquellas bacterias incapaces de crecer usando materia orgánica como fuente de carbono. Mixotrofas son aquellas bacterias con metabolismo energético litotrofo (obtienen energía de compuestos inorgánicos), pero requieren sustancias orgánicas como nutrientes para su metabolismo biosintético.")
4
Catabolismo y anabolismo: papel de la obtención de energía en vincular estos procesos
Anabolismo y catabolismo. Observa que la energía obtenida durante el catabolismo (en los heterotrofos) se usa para la biosíntesis de macromoléculas (anabolismo).
 se usa para la biosíntesis de macromoléculas (anabolismo).")
5
Quimiorganotrofos respiradores: fuente de C suministra los electrones y es origen del material celular En este esquema se muestra que los quimiorganotrofos respiradores hacen dos cosas con la fuente de C: obtener electrones para sacar energía, y derivar parte del C para los procesos biosintéticos. El aceptor de electrones al final de la c.t.e. puede ser oxígeno (respiración aerobia) u otro aceptor (inorgánico u orgánico) respiración anaerobia.
 u otro aceptor (inorgánico u orgánico) respiración anaerobia.")
6
Quimiolitotrofos: fuente de electrones inorgánica
Quimiolitotrofos: fuente de electrones inorgánica. El C celular viene del CO2 (autotrofía) Los quimiolitrofos son respiradores, pero como su fuente exógena de electrones es inorgánica, no pueden usarla para derivar material celular, sino solamente energía. El C celular lo derivan de fijación del CO2 (litotrofía).
 Los quimiolitrofos son respiradores, pero como su fuente exógena de electrones es inorgánica, no pueden usarla para derivar material celular, sino solamente energía. El C celular lo derivan de fijación del CO2 (litotrofía).")
7
Requerimiento de nutrientes comunes
Todas las bacterias necesitan captar elementos químicos, que según las cantidades en que son requeridos se clasifican en: En la naturaleza, estos elementos se encuentran combinados, formando parte de sustancias orgánicas o inorgánicas. Algunos serán incorporados para construir macromoléculas y estructuras celulares; otros solo sirven para la producción de energía; finalmente, otros pueden ejercer ambos papeles. Macronutrientes: C, H, O, N, P, S, K, Mg, Ca, Fe Micronutrientes (trazas): Mn, Co, Cu, Zn, Mo, Ni, etc. Todas las bacterias necesitan captar una serie de elementos químicos, que se pueden clasificar (según las cantidades en que son requeridos) como macronutrientes (C, H, O, N, P, S, K, Mg), y micronutrientes o elementos traza (Co, Cu, Zn, Mo...). En la naturaleza, estos elementos se encuentran combinados, formando parte de sustancias orgánicas o inorgánicas. Algunos de los nutrientes serán incorporados para construir macromoléculas y estructuras celulares; otros solo sirven para la producción de energía, y no se incorporan directamente como material celular; finalmente, otros pueden ejercer ambos papeles. El mundo bacteriano, como conjunto, exhibe una gigantesca versatilidad metabólica de uso de nutrientes: desde autotrofos que obtienen su carbono por reducción del CO2 y los demás elementos a partir de fuentes igualmente inorgánicas, hasta heterotrofos capaces de usar amplia gama de fuentes orgánicas de carbono. A su vez, dentro de los heterotrofos, podemos encontrar muchos y variados tipos de nutrición, desde bacterias metilotrofas que sólo usan metano o metanol como fuente de carbono y energía, hasta los muy versátiles Pseudomonas, que pueden recurrir a degradar más de 100 tipos de fuentes de C (incluyendo entre ellas sustancias tan “exóticas” como hidrocarburos alifáticos y cíclicos). De cualquier modo, entre los heterotrofos, una de las fuentes más típicas de carbono consiste en glucosa. En los heterotrofos-organotrofos, los sustratos carbonados (con un nivel de oxidación no muy distinto del material celular -CH2O-) entran simultáneamente a metabolismo energético (donde la fuente de C se transforma en CO2, o en CO2 junto con otras sustancias no totalmente oxidadas) y a metabolismo plástico.
: Mn, Co, Cu, Zn, Mo, Ni, etc. Todas las bacterias necesitan captar una serie de elementos químicos, que se pueden clasificar (según las cantidades en que son requeridos) como macronutrientes (C, H, O, N, P, S, K, Mg), y micronutrientes o elementos traza (Co, Cu, Zn, Mo...). En la naturaleza, estos elementos se encuentran combinados, formando parte de sustancias orgánicas o inorgánicas. Algunos de los nutrientes serán incorporados para construir macromoléculas y estructuras celulares; otros solo sirven para la producción de energía, y no se incorporan directamente como material celular; finalmente, otros pueden ejercer ambos papeles. El mundo bacteriano, como conjunto, exhibe una gigantesca versatilidad metabólica de uso de nutrientes: desde autotrofos que obtienen su carbono por reducción del CO2 y los demás elementos a partir de fuentes igualmente inorgánicas, hasta heterotrofos capaces de usar amplia gama de fuentes orgánicas de carbono. A su vez, dentro de los heterotrofos, podemos encontrar muchos y variados tipos de nutrición, desde bacterias metilotrofas que sólo usan metano o metanol como fuente de carbono y energía, hasta los muy versátiles Pseudomonas, que pueden recurrir a degradar más de 100 tipos de fuentes de C (incluyendo entre ellas sustancias tan exóticas como hidrocarburos alifáticos y cíclicos). De cualquier modo, entre los heterotrofos, una de las fuentes más típicas de carbono consiste en glucosa. En los heterotrofos-organotrofos, los sustratos carbonados (con un nivel de oxidación no muy distinto del material celular -CH2O-) entran simultáneamente a metabolismo energético (donde la fuente de C se transforma en CO2, o en CO2 junto con otras sustancias no totalmente oxidadas) y a metabolismo plástico.")
8
Diversidad metabólica de los microorganismos
El mundo microbiano es de una sorprendente diversidad metabólica. Algunos metabolismos solo han evolucionado en procariotas. Ejemplos: En heterotrofos: desde metilotrofos (usan metano o metanol) hasta los versátiles Pseudomonas, que usan más de 100 tipos de C orgánico, incluyendo hidrocarburos alifáticos y cíclicos Los quimiolitoautotrofos crecen en oscuridad en medios a base solamente de sales minerales La fijación de N2 solo ha evolucionado en procariotas
 hasta los versátiles Pseudomonas, que usan más de 100 tipos de C orgánico, incluyendo hidrocarburos alifáticos y cíclicos. Los quimiolitoautotrofos crecen en oscuridad en medios a base solamente de sales minerales. La fijación de N2 solo ha evolucionado en procariotas.")
9
Clases de nutrientes Universales (los requeridos en esta forma por todos los procariotas): H2O, CO2, fosfatos y sales minerales Particulares: elementos que se pueden captar de diferentes maneras, según especies: N y S. Especiales: los microorganismos pueden tener necesidades especiales. Ej: Las diatomeas necesitan ácido silícico Las bacterias halófilas requieren grandes cantidades de Na (mares) Factores de crecimiento Podemos clasificar los nutrientes en las siguientes categorías: · Universales (es decir, aquellos que son requeridos por todos los procariotas): agua, CO2, fosfatos y sales minerales; · Particulares; · Factores de crecimiento.
 Factores de crecimiento. Podemos clasificar los nutrientes en las siguientes categorías: · Universales (es decir, aquellos que son requeridos por todos los procariotas): agua, CO2, fosfatos y sales minerales; · Particulares; · Factores de crecimiento.")
10
El agua El agua es: Fuentes de agua:
el principal constituyente del protoplasto bacteriano; el medio universal donde ocurren las reacciones biológicas; un reactante en exceso (es decir, un producto resultante de algunas reacciones bioquímicas). Fuentes de agua: endógena (procedente de oxido-reducciones) exógena (la mayoría) procedente del medio, y que difunde a través de las membranas. Las bacterias necesitan grandes cantidades de agua. De hecho, salvo excepciones, se pueden considerar como organismos acuáticos. Requieren cierto grado de humedad para crecer. Desde el punto de vista de sus posibles papeles, el agua es: el principal constituyente del protoplasto bacteriano; el medio universal donde ocurren las reacciones biológicas; un reactante en exceso (es decir, un producto resultante de algunas reacciones bioquímicas). Las fuentes de agua pueden ser: endógena: procedente de procesos de oxido-reducción; exógena (la más importante): procedente del medio, y que difunde a través de las membranas. Ahora bien, no toda el agua de un ambiente está disponible para la bacteria: Existen determinadas sustancias que absorben y superficies que adsorben de modo más o menos intenso moléculas de agua, dejándolas inasequibles para la bacteria. Los solutos disueltos en agua (p. ej., sales, azúcares) tienen afinidad por las moléculas de H2O que los rodean, por lo que éstas tampoco estarán a disposición del microorganismo. La disponibilidad de agua se mide por un parámetro llamado actividad de agua o potencial de agua, indicativo del agua libre, y que se expresa como aW= PS/PW .donde PS es la presión parcial de vapor de agua en la solución problema y PW es la presión parcial de vapor del agua destilada. Las bacterias tienen valores de aW normalmente entre 0.90 y 0.99, auque bacterias marinas como ciertos Vibrio y Pseudomonas encuentran valores de El extremo (0.75) lo tenemos en la arquea Halobacterium, que habita en lagunas hipersalinas.
. Fuentes de agua: endógena (procedente de oxido-reducciones) exógena (la mayoría) procedente del medio, y que difunde a través de las membranas. Las bacterias necesitan grandes cantidades de agua. De hecho, salvo excepciones, se pueden considerar como organismos acuáticos. Requieren cierto grado de humedad para crecer. Desde el punto de vista de sus posibles papeles, el agua es: el principal constituyente del protoplasto bacteriano; el medio universal donde ocurren las reacciones biológicas; un reactante en exceso (es decir, un producto resultante de algunas reacciones bioquímicas). Las fuentes de agua pueden ser: endógena: procedente de procesos de oxido-reducción; exógena (la más importante): procedente del medio, y que difunde a través de las membranas. Ahora bien, no toda el agua de un ambiente está disponible para la bacteria: Existen determinadas sustancias que absorben y superficies que adsorben de modo más o menos intenso moléculas de agua, dejándolas inasequibles para la bacteria. Los solutos disueltos en agua (p. ej., sales, azúcares) tienen afinidad por las moléculas de H2O que los rodean, por lo que éstas tampoco estarán a disposición del microorganismo. La disponibilidad de agua se mide por un parámetro llamado actividad de agua o potencial de agua, indicativo del agua libre, y que se expresa como aW= PS/PW .donde PS es la presión parcial de vapor de agua en la solución problema y PW es la presión parcial de vapor del agua destilada. Las bacterias tienen valores de aW normalmente entre 0.90 y 0.99, auque bacterias marinas como ciertos Vibrio y Pseudomonas encuentran valores de El extremo (0.75) lo tenemos en la arquea Halobacterium, que habita en lagunas hipersalinas.")
11
El agua La disponibilidad de agua se mide como actividad de agua (potencial de agua; aw) aW= PS/PW . Donde PS es la presión parcial de vapor de agua en la solución problema y PW es la presión parcial de vapor del agua destilada. Las bacterias tienen valores de aw normalmente entre 0.90 y 0.99 en bacterias que viven en sangre y fluidos, aw = 0.995 en bacterias marinas. Vibrio, Pseudomona aw = 0.980 microorganismos xerófilos (aw en torno a 0.75) arqueas halófilas extremas (Halobacterium) levaduras sacarófilas, que viven en zumos y jugos
 arqueas halófilas extremas (Halobacterium) levaduras sacarófilas, que viven en zumos y jugos.")
12
El Carbono El C es necesario para construir el esqueleto de todas las moléculas orgánicas Autótrofos: pueden usar CO2 como única fuente de C Heterótrofos: emplean moléculas orgánicas preformadas y reducidas como fuente de C Los microorganismos tienen gran flexibilidad con respecto a la fuente de C

13
Fijación del CO2 La reducción o incorporación del CO2 requiere una gran cantidad de energía. Casi todos los autótrofos microbianos incorporan CO2 mediante el Ciclo de Calvin. Tiene lugar en los cloroplastos (eucariotas) o carboxisomas (procariotas). La formación de glucosa a partir de CO2 puede resumirse: 6CO ATP +12NADPH + 12H+ + 12H2O glucosa + 18(ADP+Pi) + 12NADP+ Los azúcares formados pueden usarse para sintetizar otras moléculas esenciales
 o carboxisomas (procariotas). La formación de glucosa a partir de CO2 puede resumirse: 6CO ATP +12NADPH + 12H+ + 12H2O glucosa + 18(ADP+Pi) + 12NADP+ Los azúcares formados pueden usarse para sintetizar otras moléculas esenciales.")
14
El CO2 El CO2 es requerido por todo tipo de bacterias.
Los autotrofos lo requieren como fuente de C, y lo reducen usando como fuente de energía sustancias químicas : quimioautotrofos la luz: fotoautotrofos Las arqueas metanogénicas lo pueden usar como aceptor final de electrones en la respiración, produciendo CH4. Además, algunas lo usan también como fuente de C Los heterotrofos necesitan pequeñas cantidades de CO2 para sus carboxilaciones en rutas metabólicas El anhidrido carbónico es requerido por todo tipo de bacterias: Las autotrofas lo requieren como fuente de carbono, y lo reducen usando como fuente de energía la luz (en el caso de las fotoautotrofas) u oxidaciones de determinadas sustancias inorgánicas (los quimioautolitotrofos). Las arqueas metanogénicas pueden usar el CO2 como aceptor de los electrones procedentes de la oxidación del H2, proceso por el que obtienen su energía de modo litotrofo: CO2 + 4H2 CH4 + 2H2O (G'0<0) Además, algunas arqueas no sólo usan CO2 como aceptor de electrones para obtener energía, sino que, además lo usan como fuente de carbono celular (arqueas metanogénicas autotrofas). Los heterotrofos, aunque no usan el CO2 como fuente de C ni como aceptor de electrones, necesitan pequeñas cantidades para las carboxilaciones en determinadas rutas anabólicas y catabólicas. El origen del CO2 puede ser: endógeno: procedente de descarboxilaciones que ocurren al degradar la fuente orgánica de carbono; exógeno: el CO2 de la atmósfera o disuelto en las soluciones acuosas. Normalmente, las bacterias crecen a la concentración de CO2 atmosférico (0.03%), pero algunas bacterias (Neisseria, Brucella), cuando se aislan por primera vez, requieren atmósferas enriquecidas, con 5-10% de CO2. Ello parece deberse a que poseen alguna enzima con baja afinidad hacia el carbónico; sin embargo, tras varios subcultivos, suelen adaptarse a crecer a tensiones normales.
 u oxidaciones de determinadas sustancias inorgánicas (los quimioautolitotrofos). Las arqueas metanogénicas pueden usar el CO2 como aceptor de los electrones procedentes de la oxidación del H2, proceso por el que obtienen su energía de modo litotrofo: CO2 + 4H2 CH4 + 2H2O (G 0<0) Además, algunas arqueas no sólo usan CO2 como aceptor de electrones para obtener energía, sino que, además lo usan como fuente de carbono celular (arqueas metanogénicas autotrofas). Los heterotrofos, aunque no usan el CO2 como fuente de C ni como aceptor de electrones, necesitan pequeñas cantidades para las carboxilaciones en determinadas rutas anabólicas y catabólicas. El origen del CO2 puede ser: endógeno: procedente de descarboxilaciones que ocurren al degradar la fuente orgánica de carbono; exógeno: el CO2 de la atmósfera o disuelto en las soluciones acuosas. Normalmente, las bacterias crecen a la concentración de CO2 atmosférico (0.03%), pero algunas bacterias (Neisseria, Brucella), cuando se aislan por primera vez, requieren atmósferas enriquecidas, con 5-10% de CO2. Ello parece deberse a que poseen alguna enzima con baja afinidad hacia el carbónico; sin embargo, tras varios subcultivos, suelen adaptarse a crecer a tensiones normales.")
15
El CO2 El origen del CO2 puede ser:
Endógeno: procedente de descarboxilaciones que ocurren al degradar la fuente orgánica de carbono Exógeno: el CO2 de la atmósfera o disuelto en las soluciones acuosas Normalmente, las bacterias crecen a la concentración de CO2 atmosférico (0.03%), pero algunas bacterias (Neisseria, Brucella), cuando se aislan por primera vez, requieren atmósferas enriquecidas, con 5-10% de CO2. (capnofilia o microaerofilia) El anhidrido carbónico es requerido por todo tipo de bacterias: Las autotrofas lo requieren como fuente de carbono, y lo reducen usando como fuente de energía la luz (en el caso de las fotoautotrofas) u oxidaciones de determinadas sustancias inorgánicas (los quimioautolitotrofos). Las arqueas metanogénicas pueden usar el CO2 como aceptor de los electrones procedentes de la oxidación del H2, proceso por el que obtienen su energía de modo litotrofo: CO2 + 4H2 CH4 + 2H2O (G'0<0) Además, algunas arqueas no sólo usan CO2 como aceptor de electrones para obtener energía, sino que, además lo usan como fuente de carbono celular (arqueas metanogénicas autotrofas). Los heterotrofos, aunque no usan el CO2 como fuente de C ni como aceptor de electrones, necesitan pequeñas cantidades para las carboxilaciones en determinadas rutas anabólicas y catabólicas. El origen del CO2 puede ser: endógeno:; exógeno:.
, pero algunas bacterias (Neisseria, Brucella), cuando se aislan por primera vez, requieren atmósferas enriquecidas, con 5-10% de CO2. (capnofilia o microaerofilia) El anhidrido carbónico es requerido por todo tipo de bacterias: Las autotrofas lo requieren como fuente de carbono, y lo reducen usando como fuente de energía la luz (en el caso de las fotoautotrofas) u oxidaciones de determinadas sustancias inorgánicas (los quimioautolitotrofos). Las arqueas metanogénicas pueden usar el CO2 como aceptor de los electrones procedentes de la oxidación del H2, proceso por el que obtienen su energía de modo litotrofo: CO2 + 4H2 CH4 + 2H2O (G 0<0) Además, algunas arqueas no sólo usan CO2 como aceptor de electrones para obtener energía, sino que, además lo usan como fuente de carbono celular (arqueas metanogénicas autotrofas). Los heterotrofos, aunque no usan el CO2 como fuente de C ni como aceptor de electrones, necesitan pequeñas cantidades para las carboxilaciones en determinadas rutas anabólicas y catabólicas. El origen del CO2 puede ser: endógeno:; exógeno:.")
16
Fosfatos El P suele requerirse en forma de fosfatos
Bacterias que usan fosfatos orgánicos poseen fosfatasas extracelulares (secretadas) en Gram-positivas, periplásmicas en Gram-negativas Fosfatos inorgánicos Las bacterias que usan fosfatos orgánicos no dependen de ellos, ya que también pueden usar fosfatos inorgánicos El fósforo se usa principalmente para la síntesis de los ácidos nucleicos y los fosfolípidos, pero aparece también en coenzimas y en proteínas. El fósforo suele requerirse en forma de fosfatos, sea orgánicos o inorgánicos. Las bacterias que pueden usar los fosfatos orgánicos (merced a la posesión de fosfatasas) no dependen absolutamente de ellos, ya que pueden recurrir igualmente a los fosfatos inorgánicos. Los fosfatos orgánicos son hidrolizados por fosfatasas extracelulares o (en las Gram-negativas) periplásmicas (p.ej., la fosfatasa alcalina).
 en Gram-positivas, periplásmicas en Gram-negativas. Fosfatos inorgánicos. Las bacterias que usan fosfatos orgánicos no dependen de ellos, ya que también pueden usar fosfatos inorgánicos. El fósforo se usa principalmente para la síntesis de los ácidos nucleicos y los fosfolípidos, pero aparece también en coenzimas y en proteínas. El fósforo suele requerirse en forma de fosfatos, sea orgánicos o inorgánicos. Las bacterias que pueden usar los fosfatos orgánicos (merced a la posesión de fosfatasas) no dependen absolutamente de ellos, ya que pueden recurrir igualmente a los fosfatos inorgánicos. Los fosfatos orgánicos son hidrolizados por fosfatasas extracelulares o (en las Gram-negativas) periplásmicas (p.ej., la fosfatasa alcalina).")
17
Sales minerales: cationes
Ion Potasio K+ en activación de enzimas asociado con Ac. teicoicos de Gram+ Ion Magnesio Mg2+ estabiliza ribosomas, membranas y ácidos nucleicos cofactor en reacciones con ATP en clorofilas y bacterioclorofilas Ion Calcio Ca2+ cofactor de enzimas como proteinasas El Hierro como ion Fe2+ en citocromos, FeS-proteínas cofactor en enzimas Las sales minerales son la fuente de aniones (p. ej. el Cl--) y de cationes para la célula. Los siguientes cationes, concretamente, se necesitan en cantidades relativamente grandes: K+, Mg++, Ca++, Fe++. El ión potasio (K+): interviene en la activación de una variedad de enzimas, incluyendo las que participan en la síntesis de proteínas. En Gram-positivas está asociado con los ácidos teicoicos de la pared. El ión magnesio (Mg++): estabiliza ribosomas, membranas y ácidos nucleicos; como cofactor en muchas reacciones, especialmente las que implican transferencia de grupos fosfato. Por ejemplo, en las reacciones que requieren ATP, el Mg++ puede unir la enzima al sustrato durante el mecanismo de acción de la primera. Participa de las clorofilas y bacterioclorofilas de bacterias fotosintéticas. El ión calcio (Ca++): es un cofactor de ciertas enzimas, como proteinasas. El hierro participa en muchas moléculas implicadas en procesos de respiración, como citocromos y ferroproteínas no hémicas (proteínas con Fe-S); interviene como cofactor en ciertas enzimas. Aparte de estos iones que se requieren en cantidades relativamente grandes, las bacterias necesitan minúsculas cantidades de otros elementos (oligoelementos), a los que también se denomina como micronutrientes o elementos traza: El manganeso (Mn++) es un cofactor de ciertas enzimas, y a veces puede sustituir al Mg++. El cobalto (Co++) se requiere casi exclusivamente para la vitamina B12 (de hecho, si suministramos esta vitamina al medio, la bacteria se vuelve independiente del Co++ libre). El zinc interviene en la estabilización de complejos enzimáticos como las ADN- y ARN-polimerasas. El Mo participa en las llamadas molibdoflavoproteínas, implicadas en la asimilación de nitratos. Por otro lado, participa como cofactor, junto con el Fe, en el complejo nitrogenasa de las bacterias fijadoras de N2 atmosférico. El níquel participa en hidrogenasas, enzimas que captan o liberan H2.
 y de cationes para la célula. Los siguientes cationes, concretamente, se necesitan en cantidades relativamente grandes: K+, Mg++, Ca++, Fe++. El ión potasio (K+): interviene en la activación de una variedad de enzimas, incluyendo las que participan en la síntesis de proteínas. En Gram-positivas está asociado con los ácidos teicoicos de la pared. El ión magnesio (Mg++): estabiliza ribosomas, membranas y ácidos nucleicos; como cofactor en muchas reacciones, especialmente las que implican transferencia de grupos fosfato. Por ejemplo, en las reacciones que requieren ATP, el Mg++ puede unir la enzima al sustrato durante el mecanismo de acción de la primera. Participa de las clorofilas y bacterioclorofilas de bacterias fotosintéticas. El ión calcio (Ca++): es un cofactor de ciertas enzimas, como proteinasas. El hierro participa en muchas moléculas implicadas en procesos de respiración, como citocromos y ferroproteínas no hémicas (proteínas con Fe-S); interviene como cofactor en ciertas enzimas. Aparte de estos iones que se requieren en cantidades relativamente grandes, las bacterias necesitan minúsculas cantidades de otros elementos (oligoelementos), a los que también se denomina como micronutrientes o elementos traza: El manganeso (Mn++) es un cofactor de ciertas enzimas, y a veces puede sustituir al Mg++. El cobalto (Co++) se requiere casi exclusivamente para la vitamina B12 (de hecho, si suministramos esta vitamina al medio, la bacteria se vuelve independiente del Co++ libre). El zinc interviene en la estabilización de complejos enzimáticos como las ADN- y ARN-polimerasas. El Mo participa en las llamadas molibdoflavoproteínas, implicadas en la asimilación de nitratos. Por otro lado, participa como cofactor, junto con el Fe, en el complejo nitrogenasa de las bacterias fijadoras de N2 atmosférico. El níquel participa en hidrogenasas, enzimas que captan o liberan H2.")
18
Oligoelementos o micronutrientes
Mn2+ (cofactor de ciertas enzimas) Co2+ (vitamina B12) Zn2+ (estabiliza ADN-polimerasas y ARN-polimerasas) Mo (en molibdoflavoproteínas, en la asimilación de nitratos. Cofactor en el complejo nitrogenasa) Ni (en hidrogenasas, enzimas que captan o liberan H2)
 Co2+ (vitamina B12) Zn2+ (estabiliza ADN-polimerasas y ARN-polimerasas) Mo (en molibdoflavoproteínas, en la asimilación de nitratos. Cofactor en el complejo nitrogenasa) Ni (en hidrogenasas, enzimas que captan o liberan H2)")
19
Nitrógeno y azufre Los elementos N y S son requeridos por todos los seres vivos, se encuentran en la célula en estado reducido: El -NH2 forma parte de los aminoácidos y de las bases nitrogenadas El radical –SH interviene en aminoácidos y coenzimas Se trata de elementos que pueden ser cubiertos de modo muy distinto, dependiendo del tipo de bacteria que consideremos. Concretamente, los elementos N y S (que requieren todos los seres vivos) pueden ser captados por las bacterias de modos muy distintos, dependiendo de sus capacidades biosintéticas. Tanto el N como el S se encuentran en la célula en estado reducido: el radical -NH2 forma parte de los aminoácidos (que a su vez son los sillares de las proteínas) y de las bases nitrogenadas (que participan en los ácidos nucleicos y en algunas coenzimas); el radical -SH interviene en determinados aminoácidos y en coenzimas como la CoA. ¿En qué formas químicas entran N y S a las bacterias? La mayoría de bacterias fotosintéticas y muchas heterotrofas asimilan estos elementos en forma combinada inorgánica oxidada: como NO3--, merced a la actuación secuencial de nitrato-reductasas y nitrito-reductasas asimilatorias. Como SO42-. Este sulfato se activa con ATP, y luego se reduce hasta sulfito y finalmente sulfhídrico, que ya tiene el estado de reducción adecuado para la incorporación del S. Muchas bacterias heterotrofas pueden usar alguna forma reducida: de N inorgánico: amonio (NH4+); de S inorgánico: sulfuros (S2-, SH-); de N orgánico: aminoácidos, péptidos; de S orgánico:cisteína. Muchas de las bacterias que pueden usar amonio como única fuente de nitrógeno también pueden usar nitratos.
 pueden ser captados por las bacterias de modos muy distintos, dependiendo de sus capacidades biosintéticas. Tanto el N como el S se encuentran en la célula en estado reducido: el radical -NH2 forma parte de los aminoácidos (que a su vez son los sillares de las proteínas) y de las bases nitrogenadas (que participan en los ácidos nucleicos y en algunas coenzimas); el radical -SH interviene en determinados aminoácidos y en coenzimas como la CoA. ¿En qué formas químicas entran N y S a las bacterias La mayoría de bacterias fotosintéticas y muchas heterotrofas asimilan estos elementos en forma combinada inorgánica oxidada: como NO3--, merced a la actuación secuencial de nitrato-reductasas y nitrito-reductasas asimilatorias. Como SO42-. Este sulfato se activa con ATP, y luego se reduce hasta sulfito y finalmente sulfhídrico, que ya tiene el estado de reducción adecuado para la incorporación del S. Muchas bacterias heterotrofas pueden usar alguna forma reducida: de N inorgánico: amonio (NH4+); de S inorgánico: sulfuros (S2-, SH-); de N orgánico: aminoácidos, péptidos; de S orgánico:cisteína. Muchas de las bacterias que pueden usar amonio como única fuente de nitrógeno también pueden usar nitratos.")
20
Captación de N y S El N y el S pueden ser captados de modos distintos, según capacidades biosintéticas. En forma combinada inorgánica oxidada: NO3- (acción de nitratorreductasas y nitritorreductasas asimilatorias) NH3 N orgánico SO42- (se activa con ATP, y luego se reduce a SO32- y finalmente hasta SH2 entra a comp. Orgánicos En forma combinada reducida N reducido inorgánico: NH4+ S reducido inorgánico: S2-, SH- N reducido orgánico: aminoácidos, péptidos S reducido orgánico: cisteína
 NH3 N orgánico. SO42- (se activa con ATP, y luego se reduce a SO32- y finalmente hasta SH2 entra a comp. Orgánicos. En forma combinada reducida. N reducido inorgánico: NH4+ S reducido inorgánico: S2-, SH- N reducido orgánico: aminoácidos, péptidos. S reducido orgánico: cisteína.")
21
N2 + 8H+ + 8e + 18 ATP 2NH3 + H2 + 18 (ADP + Pi)
Fijación de N2 La capacidad de nutrición nitrogenada a partir del N2 atmosférico (N libre, no combinado) solo ha evolucionado en ciertos procariotas: procariotas diazotrofos o fijadores de nitrógeno N2 + 8H+ + 8e ATP 2NH3 + H2 + 18(ADP+Pi) Catalizada por el complejo enzimático nitrogenasa: Comp. I (= nitrogenasa propiamente dicha): MoFe-proteína (cofactor FeMoCo) Comp. II (=nitrogenasa reductasa ): Fe-proteína La atmósfera contiene enormes cantidades de nitrógeno no combinado (libre) en estado gaseoso: el nitrógeno molecular o dinitrógeno (N2), que procede de microorganismos desnitrificantes (véase el capítulo 10). Sin embargo, esta gran reserva sólo puede servir de fuente de nitrógeno a ciertos procariotas, las llamadas bacterias fijadoras de nitrógeno o diazotrofos. Esta notable capacidad bioquímica no ha evolucionado en eucariotas. (Lo más a que ha llegado la evolución es a seleccionar ciertos tipos de asociaciones simbióticas entre procariotas diazotrofos y ciertos eucariotas, como p. ej., la simbiosis entre raíces de leguminosas y bacterias del grupo de Rhizobium). La fijación del N2 es un proceso de reducción que convierte el nitrógeno molecular en amoniaco, según la siguiente ecuación: N2 + 8H+ + 8e ATP 2NH3 + H (ADP + Pi) Esta reacción está catalizada por un complejo enzimático denominado nitrogenasa o dinitrogenasa, que consta de dos componentes: · Componente I o nitrogenasa propiamente dicha; posee un cofactor de hierro y molibdeno (FeMoCo) que forma parte del centro activo. Por ello, a este componente también se le conoce como molibdoferroproteína. (En realidad existen dos copias del cofactor, cuya estequiometría es MoFe7S8-homocitrato) · Componente II o nitrogenasa-reductasa, que posee átomos de Fe acomplejados con S de determinadas cisteínas (por lo que este componente se denomina a veces ferroproteína).
 solo ha evolucionado en ciertos procariotas: procariotas diazotrofos o fijadores de nitrógeno. N2 + 8H+ + 8e ATP 2NH3 + H2 + 18(ADP+Pi) Catalizada por el complejo enzimático nitrogenasa: Comp. I (= nitrogenasa propiamente dicha): MoFe-proteína (cofactor FeMoCo) Comp. II (=nitrogenasa reductasa ): Fe-proteína. La atmósfera contiene enormes cantidades de nitrógeno no combinado (libre) en estado gaseoso: el nitrógeno molecular o dinitrógeno (N2), que procede de microorganismos desnitrificantes (véase el capítulo 10). Sin embargo, esta gran reserva sólo puede servir de fuente de nitrógeno a ciertos procariotas, las llamadas bacterias fijadoras de nitrógeno o diazotrofos. Esta notable capacidad bioquímica no ha evolucionado en eucariotas. (Lo más a que ha llegado la evolución es a seleccionar ciertos tipos de asociaciones simbióticas entre procariotas diazotrofos y ciertos eucariotas, como p. ej., la simbiosis entre raíces de leguminosas y bacterias del grupo de Rhizobium). La fijación del N2 es un proceso de reducción que convierte el nitrógeno molecular en amoniaco, según la siguiente ecuación: N2 + 8H+ + 8e + 18 ATP 2NH3 + H (ADP + Pi) Esta reacción está catalizada por un complejo enzimático denominado nitrogenasa o dinitrogenasa, que consta de dos componentes: · Componente I o nitrogenasa propiamente dicha; posee un cofactor de hierro y molibdeno (FeMoCo) que forma parte del centro activo. Por ello, a este componente también se le conoce como molibdoferroproteína. (En realidad existen dos copias del cofactor, cuya estequiometría es MoFe7S8-homocitrato) · Componente II o nitrogenasa-reductasa, que posee átomos de Fe acomplejados con S de determinadas cisteínas (por lo que este componente se denomina a veces ferroproteína).")
22
Mecanismo de la fijación de N2
Los electrones llegan al complejo por medio de una FeS-proteína no hémica (ferredoxina o flavodoxina) Los e- se transfieren a la nitrogenasa reductasa se reduce, liga 2 ATP y se une a la dinitrogenasa La dinitrogenasa se reduce, los ATP se hidrolizan, mientras la nitrogenasa reductasa se disocia y queda preparada para otra ronda de transferencia de electrones El dinitrogenasa reducida cede (a través de su FeMoCo) electrones al N2 y junto con protones NH3 Los electrones para la reducción llegan al complejo por medio de una ferredoxina o una flavodixina (FeS proteínas no hémicas), que los transfiere al componente II, que queda reducido. El componente II reducido se une a dos moléculas de ATP, y cambia su conformación, lo que le permite unirse al componente I. Entonces se produce la transferencia de electrones desde el componente II al componente I, con hidrólisis de ATP, lo que a su vez provoca la separación del componente II respecto del I. Una vez reducido el componente I (la molibdoferroproteína), éste transfiere los electrones (y los protones) al N2, hasta convertirlo en dos moléculas de amoniaco. (El centro activo es FeMoCo). El amoniaco entra entonces en las rutas biosintéticas para convertirlo en N orgánico, incorporable a las macromoléculas.
 Los e- se transfieren a la nitrogenasa reductasa se reduce, liga 2 ATP y se une a la dinitrogenasa. La dinitrogenasa se reduce, los ATP se hidrolizan, mientras la nitrogenasa reductasa se disocia y queda preparada para otra ronda de transferencia de electrones. El dinitrogenasa reducida cede (a través de su FeMoCo) electrones al N2 y junto con protones NH3. Los electrones para la reducción llegan al complejo por medio de una ferredoxina o una flavodixina (FeS proteínas no hémicas), que los transfiere al componente II, que queda reducido. El componente II reducido se une a dos moléculas de ATP, y cambia su conformación, lo que le permite unirse al componente I. Entonces se produce la transferencia de electrones desde el componente II al componente I, con hidrólisis de ATP, lo que a su vez provoca la separación del componente II respecto del I. Una vez reducido el componente I (la molibdoferroproteína), éste transfiere los electrones (y los protones) al N2, hasta convertirlo en dos moléculas de amoniaco. (El centro activo es FeMoCo). El amoniaco entra entonces en las rutas biosintéticas para convertirlo en N orgánico, incorporable a las macromoléculas.")
23
Mecanismo de acción del complejo nitrogenasa
Mecanismo de la nitrogenasa (ver nota de la diapositiva anterior, para más detalles). Observa que, aparte de N2, la nitrogenasa puede usar otros sustratos para su reducción: acetileno y protones.
. Observa que, aparte de N2, la nitrogenasa puede usar otros sustratos para su reducción: acetileno y protones.")
24
Peculiaridades de la fijación de nitrógeno
Grandes exigencias energéticas (al menos 18 ATP), debido a que el triple enlace NN tiene una alta energía de disociación (N2 es muy inerte) requiere mucha energía activación N2 + 8H+ + 8e ATP 2NH3 + H2 + 18(ADP+Pi) Extrema sensibilidad de la nitrogenasa al oxígeno rápida e irreversible inactivación El N combinado (nitratos, amonio, aminoácidos) provoca: inhibición de la actividad nitrogenasa represión de la transcripción de los genes de fijación (nif) Dos cosas llaman la atención de la reacción de la nitrogenasa: · Obsérvese que la reacción requiere un gran aporte de energía en forma de al menos 18 ATP (en ocasiones puede llegar a 24 ATP). Ello se debe a que el dinitrógeno (NN) es una molécula extremadamente inerte (su energía de disociación es de 940 kJ), y su reducción precisa una gran energía de activación para transferirle 6 electrones. · Parte de la actividad nitrogenasa (así como ATP y electrones) “se pierden” en reducir dos iones H+ hasta H2. Se desconoce la razón de este “despilfarro”, pero se sabe que es un efecto intrínseco de este complejo enzimático. El complejo nitrogenasa tiene una extrema sensibilidad al oxígeno, de modo que queda rápida e irreversiblemente inactivado por este gas. Ahora bien, esto no significa que la capacidad de diazotrofía esté relegada a bacterias anaerobias, ya que, como veremos en la sección de taxonomía, la evolución ha “inventado” distintas estrategias para proteger a la nitrogenasa en fijadores aerobios. Debido a lo “caro” que resulta fijar nitrógeno, no es extraño comprobar que este proceso esté regulado de forma muy estricta ante la presencia en el medio de fuentes combinadas de nitrógeno (nitratos, amonio, aminoácidos): · la actividad nitrogenasa se ve inhibida ante la presencia de N combinado (nitratos, amonio, aminoácidos); · ante N combinado, se reprime la transcripción de los genes codificadores de la nitrogenasa y demás funciones relacionadas (genes nif).
, debido a que el triple enlace NN tiene una alta energía de disociación (N2 es muy inerte) requiere mucha energía activación. N2 + 8H+ + 8e ATP 2NH3 + H2 + 18(ADP+Pi) Extrema sensibilidad de la nitrogenasa al oxígeno rápida e irreversible inactivación. El N combinado (nitratos, amonio, aminoácidos) provoca: inhibición de la actividad nitrogenasa. represión de la transcripción de los genes de fijación (nif) Dos cosas llaman la atención de la reacción de la nitrogenasa: · Obsérvese que la reacción requiere un gran aporte de energía en forma de al menos 18 ATP (en ocasiones puede llegar a 24 ATP). Ello se debe a que el dinitrógeno (NN) es una molécula extremadamente inerte (su energía de disociación es de 940 kJ), y su reducción precisa una gran energía de activación para transferirle 6 electrones. · Parte de la actividad nitrogenasa (así como ATP y electrones) se pierden en reducir dos iones H+ hasta H2. Se desconoce la razón de este despilfarro , pero se sabe que es un efecto intrínseco de este complejo enzimático. El complejo nitrogenasa tiene una extrema sensibilidad al oxígeno, de modo que queda rápida e irreversiblemente inactivado por este gas. Ahora bien, esto no significa que la capacidad de diazotrofía esté relegada a bacterias anaerobias, ya que, como veremos en la sección de taxonomía, la evolución ha inventado distintas estrategias para proteger a la nitrogenasa en fijadores aerobios. Debido a lo caro que resulta fijar nitrógeno, no es extraño comprobar que este proceso esté regulado de forma muy estricta ante la presencia en el medio de fuentes combinadas de nitrógeno (nitratos, amonio, aminoácidos): · la actividad nitrogenasa se ve inhibida ante la presencia de N combinado (nitratos, amonio, aminoácidos); · ante N combinado, se reprime la transcripción de los genes codificadores de la nitrogenasa y demás funciones relacionadas (genes nif).")
25
Rhizobium - simbiosis con leguminosas

26
Factores de crecimiento
Son moléculas orgánicas, componentes celulares esenciales, que no pueden ser sintetizados por el microorganismo, los toman del ambiente, y son requeridos en muy pequeñas cantidades Existen 3 clases principales Aminoácidos: síntesis de proteínas Purinas y pirimidinas: síntesis de ácidos nucleicos Vitaminas: cofactores enzimáticos Ejemplos: Brucella requieren biotina, niacina, tiamina y pantoténico Haemophilus necesita hemo y piridín-nucleótidos Los factores de crecimiento son moléculas orgánicas específicas que, en muy pequeña cantidad, algunas bacterias necesitan para crecer. Salvo excepciones no tienen función plástica (no son sillares de macromoléculas) ni sirven como fuente de energía. Suelen ser coenzimas o sus precursores, vitaminas, que determinadas bacterias no pueden fabricar por sí mismas, al carecer de parte o toda una ruta biosintética. Ejemplos: las bacterias del género Brucella requieren como factores de crecimiento en sus medios de cultivo la biotina, niacina, tiamina y ácido pantoténico. Haemophilus necesita suplementos de grupos hemo y piridín-nucleótidos.
 ni sirven como fuente de energía. Suelen ser coenzimas o sus precursores, vitaminas, que determinadas bacterias no pueden fabricar por sí mismas, al carecer de parte o toda una ruta biosintética. Ejemplos: las bacterias del género Brucella requieren como factores de crecimiento en sus medios de cultivo la biotina, niacina, tiamina y ácido pantoténico. Haemophilus necesita suplementos de grupos hemo y piridín-nucleótidos.")
27
Factores de crecimiento
Factor o vitamina funciones principales p-aminobenzoico (PABA) precursor del ácido fólico Acido fólico metabolismo de compuestos C1, transferencia de grupos metilo Biotina biosíntesis de ácidos grasos; fijación de CO2 Cobalamina (vitamina B12) reducción y transferencia de compuestos C1; síntesis de desoxirribosa Niacina (ácido nicotínico) precursor del NAD; transferencia de electrones en reacciones redox Riboflavina precursor de FAD y FMN ácido pantoténico precursor de la CoA Tiamina (vitamina B1) descarboxilaciones; transcetolasas. Complejo B6 (piridoxal, piridoxamina) transformaciones de aminoácidos y cetoácidos Grupo Vitamina K, quinonas transportadores de electrones (ubiquinonas, menaquinonas, etc.)
 precursor del ácido fólico. Acido fólico. metabolismo de compuestos C1, transferencia de grupos metilo. Biotina. biosíntesis de ácidos grasos; fijación de CO2. Cobalamina (vitamina B12) reducción y transferencia de compuestos C1; síntesis de desoxirribosa. Niacina (ácido nicotínico) precursor del NAD; transferencia de electrones en reacciones redox. Riboflavina. precursor de FAD y FMN ácido pantoténico. precursor de la CoA. Tiamina (vitamina B1) descarboxilaciones; transcetolasas. Complejo B6 (piridoxal, piridoxamina) transformaciones de aminoácidos y cetoácidos. Grupo Vitamina K, quinonas. transportadores de electrones (ubiquinonas, menaquinonas, etc.)")
28
Prototrofos y Auxotrofos
PROTOTROFO: Un microorganismo que precisa de los mismos nutrientes que la mayoría de los miembros de su especie AUXOTROFO: Un microorganismo mutado que carece de la capacidad para sintetizar un nutriente esencial, y por ello debe obtenerlo (o a su precursor) del ambiente.
 del ambiente.")
29
Captación celular de nutrientes
Los microorganismos utilizan varios sistemas de transporte diferentes. Los más importantes son: Difusión simple Difusión facilitada Transporte activo Traslocación de grupo

30
Difusión simple Depende de la diferencia de gradiente de concentración entre el exterior y el interior de la célula, y de la permeabilidad de la membrana La concentración externa del nutriente debe ser alta Solo moléculas pequeñas pueden atravesar la membrana: H2O, O2, CO2, glicerol.

31
Difusión facilitada Intervienen proteínas transportadoras, integradas a la membrana: permeasas Las permeasas tienen afinidad específica por la sustancia que transportan. El movimiento depende del gradiente de concentración sin gasto de energía El proceso es reversible

32
Transporte activo Permite el transporte de moléculas en contra de un gradiente de concentración, con gasto de energía El complejo proteico de membrana tiene varias subunidades: poro La fuente de energía puede ser: ATP, compuestos de fosfato de alta energía, fuerza protón motriz.

33
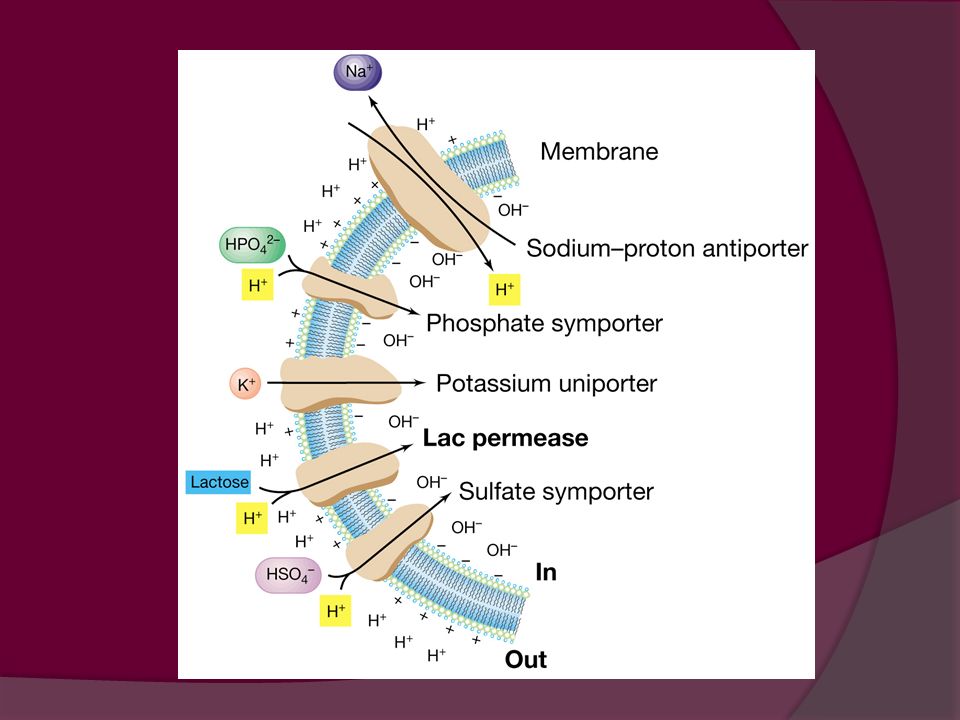
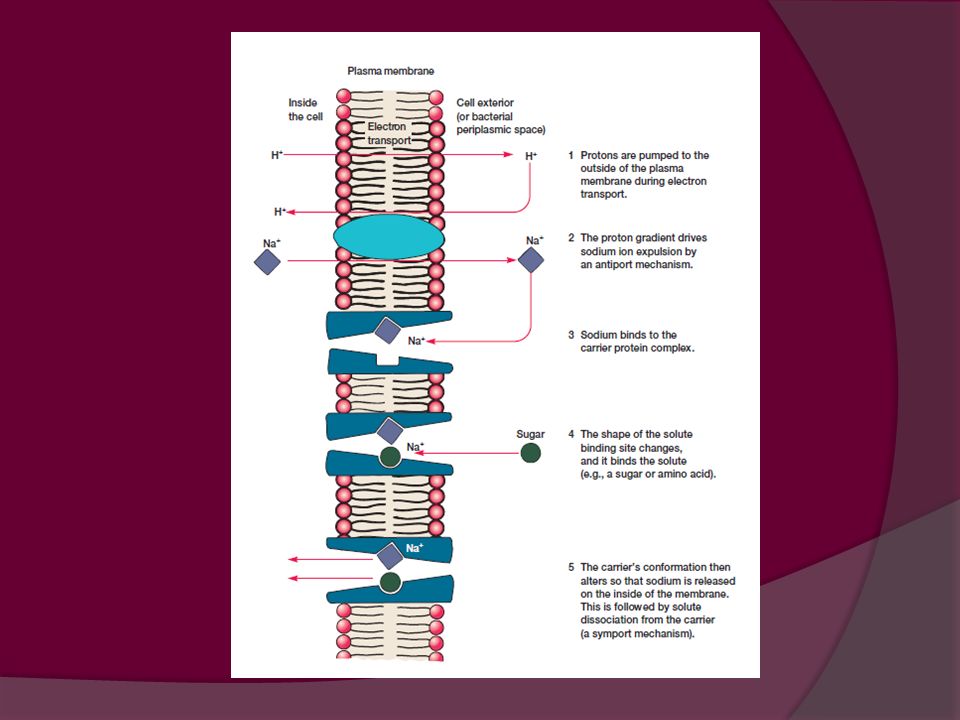
Transporte por gradiente de H+
SIMPORTE: transporte combinado de dos sustancias en la misma dirección Ej. Simporte de H+: La energía almacenada en el protón facilita el transporte del soluto. Se utiliza para captar aminoácidos y ácidos orgánicos ANTIPORTE: Sistema combinado de transporte por el que las sustancias se desplazan en direcciones opuestas. Ej. Como respuesta al ingreso de H+ se bombean iones Na+ al exterior. El gradiente de sodio generado dirige la captación de azúcares y aminoácidos

36
Traslocación de grupo Una molécula es transportada al interior celular después de alterarse químicamente Ej. Sistema Fosfoenolpiruvato-Fosfotransferasa de azúcares PEP + azúcar (exterior) / piruvato + azúcar-P (interior)
 / piruvato + azúcar-P (interior)")
37
Sistema PEP - PTS

38
Captación de Fe La captación de Fe es difícil debido a la insolubilidad del ion Fe3+ Los sideróforos son moléculas de bajo PM capaces de formar complejos de Fe3+ y aportarlos a la célula Los mo secretan sideróforos cuando hay poco Fe disponible. E complejo Fe-siderof se une a la prot receptora del sideróforo. Dentro de la célula el Fe se reduce a Fe2+

Presentaciones similares














: Autótrofos: A partir de la materia inorgánica y del CO2 de la atmósfera.>")




