Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Enzimas Q.B.P. Karla Díaz

2
Enzimas Son catalizadores de gran eficiencia y especificidad.
Aceleran la aproximación de una reacción a su equilibrio. Son muy específicas para los reactantes o sustratos, sobre los cuales actúan. El grado de especificidad del sustrato varía.

3
Las enzimas son proteínas, o bien proteínas + cofactores.
Ciertas moléculas de RNA también presentan actividad enzimática. Las moléculas de sustrato se unen a la enzima en su sitio activo. El sitio activo es una hendidura un poco hidrofóbica.

4
Nomenclatura Las enzimas reciben su nombre de acuerdo a la reacción que catalizan. Su nombre se forma añadiendo el sufijo –asa al nombre del sustrato sobre el que actúan o bien al término que describe las reacciones que catalizan. Algunas enzimas como la tripsina y la quimotripsina aún se conocen por sus nombres históricos.

5
Clasificación Un comité de la Unión Internacional de Bioquímica publicó un esquema de clasificación que asigna un número a cada enzima y clasifica a las enzimas en 6 grupos de acuerdo a la clase general de reacciones químicas que catalizan.

6
Clasificación Las óxidorreductasas catalizan reacciónes de oxidación – reducción. La mayor parte de estas enzimas se conocen como deshidrogenasas, pero algunas de ellas son oxidasas, peroxidasa, oxigenasas o reductasas. Las transferasas catalizan reacciones de transferencia de grupos. Muchas de ellas requieren la existencia de coenzimas. Estas enzimas o sus coenzimas son sustituídas en forma covalente por una porción de la molécula de sustrato. Las hidrolasas catalizan la hidrólisis. Son una clase especial de transferasas, el agua les sirve como un receptor del grupo transferido.

7
Las liasas catalizan reacciones de eliminación no hidrolítica, no oxidante, o la lisis de un sustrato en reacciones que generan un enlace doble. En dirección inversa, las liasas catalizan la adición de un sustrato a un doble enlace de un segundo sustrato. Una liasa que cataliza una reacción de adición en las células se denomina sintasa. Las isomerasas catalizan reacciones de isomerización. Debido a que estas reacciones tienen un solo sustrato y un producto, son por lo regular reacciones enzimáticas más sencillas. Las ligasas catalizan la ligadura o unión de dos sustratos en reacciones sintéticas que requieren el ingreso de la energía química potencial de un nucleósido trifosfato, como el ATP. A las ligasas se les da el nombre de sintetasas.

8
Isoenzimas Las isoenzimas son distintas formas moleculares de una misma enzima. La principal característica de las isoenzimas es que deben tener la misma especificidad de sustrato.

9
Cofactores A veces, un enzima requiere para su función la presencia de sustancias no proteicas que colaboran en la catálisis: los cofactores. Los cofactores pueden ser iones inorgánicos como el Fe++, Mg++, Mn++, Zn++ etc. Casi un tercio de los enzimas conocidos requieren cofactores. Cuando el cofactor es una molécula orgánica se llama coenzima. Muchos de estos coenzimas se sintetizan a partir de vitaminas.

10
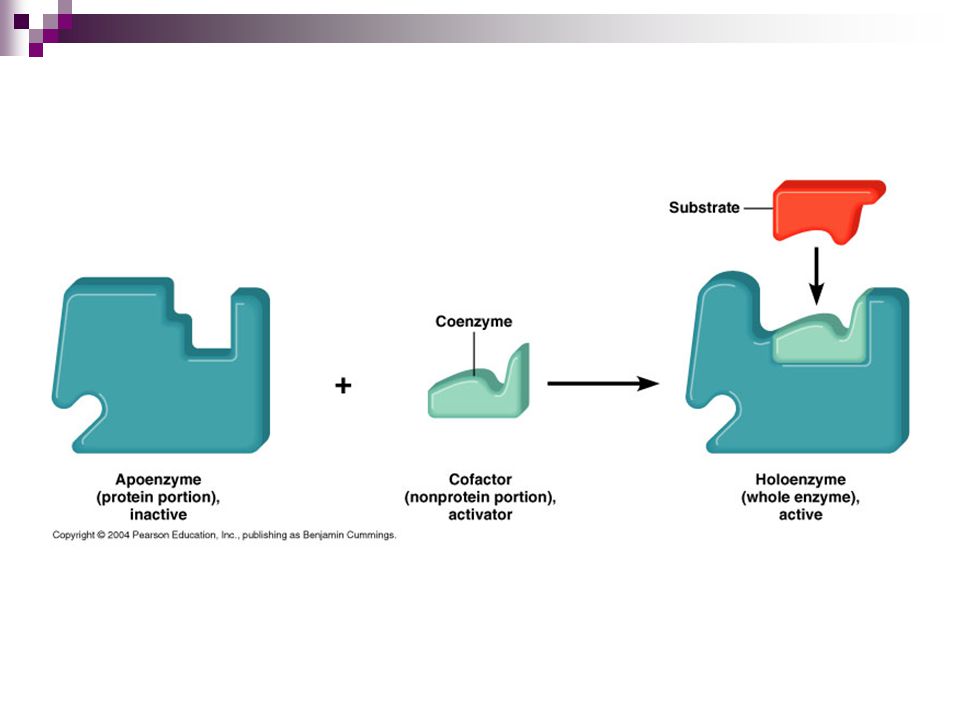
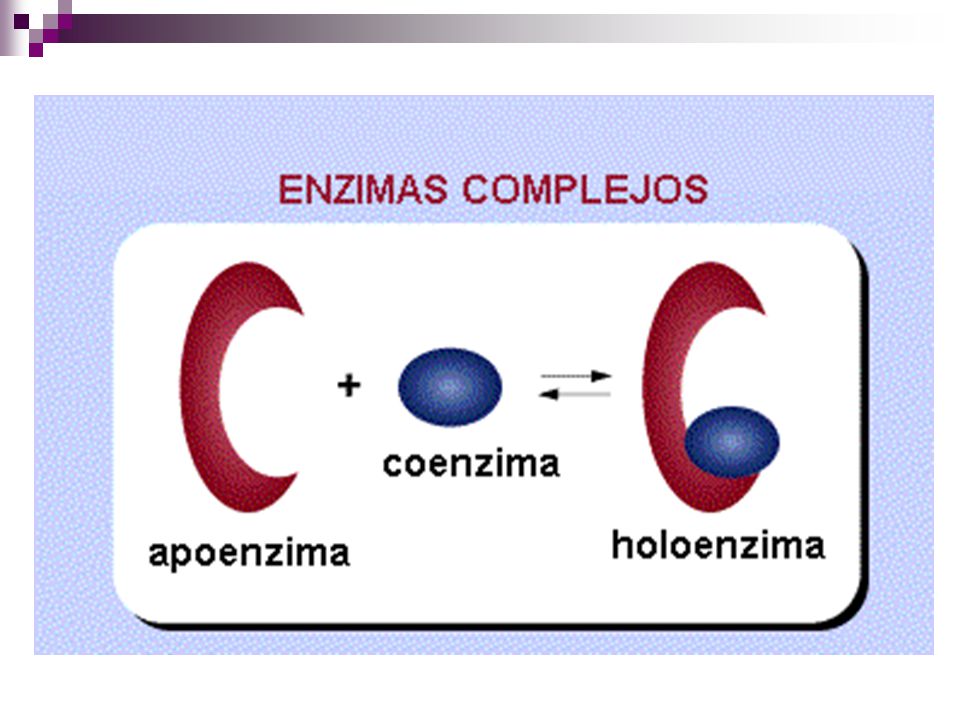
apoenzima + grupo prostético= holoenzima
Grupos prostéticos Cuando los cofactores y las coenzimas se encuentran unidos covalentemente a la enzima se llaman grupos prostéticos. La forma catalíticamente activa de la enzima, es decir, la enzima unida a su grupo prostético, se llama holoenzima. La parte proteica de una holoenzima (inactiva) se llama apoenzima, de forma que: apoenzima + grupo prostético= holoenzima
 se llama apoenzima, de forma que: apoenzima + grupo prostético= holoenzima.")
12
Cinética enzimática Michaelis-Menten

13
La tasa o velocidad de reacción de una enzima está dada por:
dS dP V = - = dt dt

14
Con respecto al tiempo…
Producto formado (mg/mL) Tiempo de reacción (min.)
 Tiempo de reacción (min.)")
15
Cinética de Michaelis-Menten
max = k2eo V (Tasa de reacción) X 103, mol/(L-sec) = max / 2 S1/2 = Km S (Concentración de sustrato) X 103, mol/L
 X 103, mol/(L-sec) = max / 2. S1/2 = Km. S (Concentración de sustrato) X 103, mol/L.")
16
Para este comportamiento enzimático, se propuso la siguiente fórmula:
maxS = max = eo Km + S = max / 2 cuando S es igual a la Km S es la concentración de sustrato libre eo es la concentración de la enzima total (libre y combinada)
")
17
Como punto inicial, se asume que la enzima E y el sustrato S se combinan para formar un complejo ES que posteriormente se disocia en el producto P y enzima libre E: K1 S + E ES K-1 K2 ES P + E

18
¨Constante de Michaelis” o de disociación.
se K-1 = = Km (es) K1
 K1.")
19
eo es la concentración total de enzima
Se asume que la descomposición del complejo ES al producto y la enzima libre es una reacción irreversible. dP = = K2 (es) dt e + (es) = eo eo es la concentración total de enzima Parámetros cinéticos max = Velocidad máxima o limitante Km = constante de Michaelis
 dt. e + (es) = eo. eo es la concentración total de enzima. Parámetros cinéticos. max = Velocidad máxima o limitante. Km = constante de Michaelis.")
20
max [S] = Km + [S] Parámetros cinéticos
max = Velocidad máxima o limitante Km = constante de Michaelis
![max [S] = Km + [S] Parámetros cinéticos](http://slideplayer.es/slide/5428410/17/images/20/%EF%81%AEmax+%5BS%5D+%EF%81%AE+%3D+Km+%2B+%5BS%5D+Par%C3%A1metros+cin%C3%A9ticos.jpg "max = Velocidad máxima o limitante. Km = constante de Michaelis.")
21
Con la expresión de Michaelis-Menten, el tiempo de reacción puede ser estimado analíticamente integrando: -max[S] dS = Km + [S] dt

22
Obteniendo: So maxt = So – S + Km ln S

23
Obtención de los parámetros cinéticos Km y Vmax
Ecuación de Lineweaver-Burk 1 1 Km 1 = + max max S Y = b + mX

24
Y = b + mX S Km 1 S = + max max Lineweaver-Burk modificada (Hanes)
Estimación de la Km sujeta a errores

25
Estimación de sujeta a errores
Eadie-Hofstee _ max = Km S Y = b + mX Estimación de sujeta a errores

26
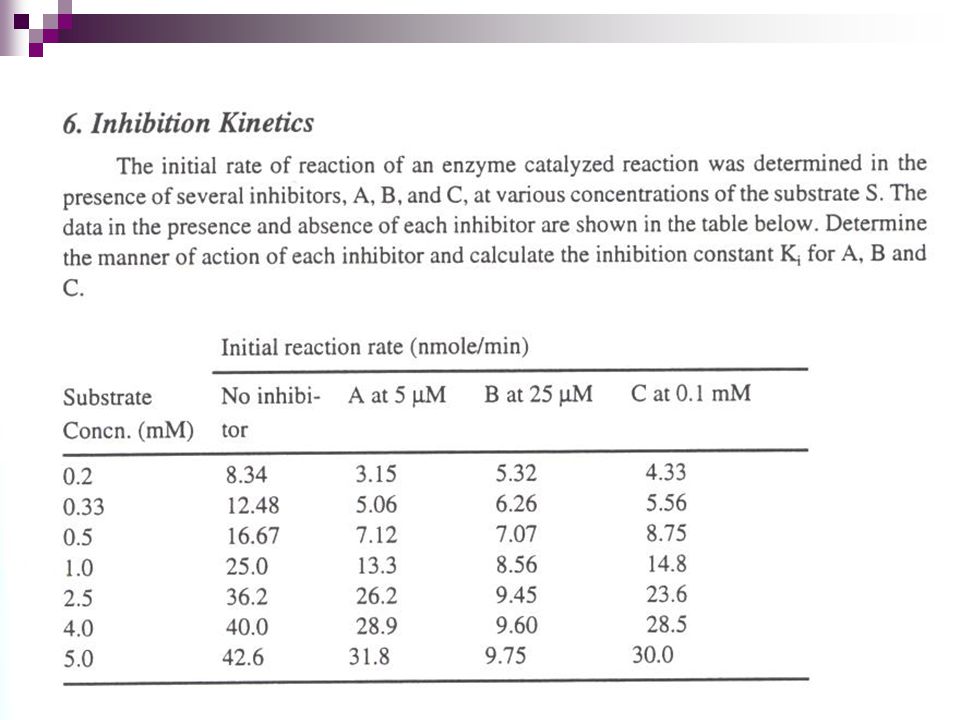
Problem. The reaction rate at different substrate concentrations for the enzyme fumarase are reported in Table 1. Using the information reported, estimate the kinetics parameters max and Km. S(g/L) Rate (g/L-min) 0.5 0.02 1.5 0.04 3.5 0.06 5.5 0.07 8.0 10.0 0.08
 Rate (g/L-min)")
27
Possible solution to the problem
S(g/L) (g/L-min) 1/S (L/g) 1/ (L-min/g) 0.5 0.02 2.0 50 1.5 0.04 0.67 23.8 3.5 0.06 0.30 17.2 5.5 0.07 0.18 14.5 8.0 10.0 0.08 0.10 12.2 Kinetics parameters max = g/L-min Km = 2.0 g/L
 (g/L-min) 1/S (L/g) 1/ (L-min/g) Kinetics parameters max = g/L-min. Km = 2.0 g/L.")
28
Inhibición enzimática

29
Un esquema general +S E + P E ES -S +I +I -I -I +S EI EIS -S

30
Inhibición competitiva
+S E + P E ES -S +I -I EI KM*=KM(1+[I]/KI)
")
31
Inhibición no competitiva
+S E + P E ES -S +I -I EIS vmax*=vmax/(1+[I]/KI)
")
32
Inhibición acompetitiva
+S E + P E ES -S +I +I -I -I +S EI EIS -S KM*=KM/(1+[I]/KI) vmax*=vmax/(1+[I]/KI)
 vmax*=vmax/(1+[I]/KI)")
33
Inhibición mixta +S E + P E ES -S +I +I -I -I +S EI EIS -S

34
¿Cómo saber que tipo de inhibición?

35
¿Cómo saber que tipo de inhibición?
KM*=KM/(1+[I]/KI) vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva Competitiva Acompetitiva
 vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva. Competitiva. Acompetitiva.")
36
¿Cómo saber que tipo de inhibición?
KM*=KM/(1+[I]/KI) vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva Competitiva Acompetitiva
 vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva. Competitiva. Acompetitiva.")
37
¿Cómo saber que tipo de inhibición?
KM*=KM/(1+[I]/KI) vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva Competitiva Acompetitiva
 vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva. Competitiva. Acompetitiva.")
38
¿Cómo saber que tipo de inhibición?
KM*=KM/(1+[I]/KI) vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva Competitiva Acompetitiva
 vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva. Competitiva. Acompetitiva.")
39
¿Cómo saber que tipo de inhibición?
KM*=KM/(1+[I]/KI) vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva Competitiva Acompetitiva
 vmax*=vmax/(1+[I]/KI) KM*=KM(1+[I]/KI) vmax*=vmax/(1+[I]/KI) No Competitiva. Competitiva. Acompetitiva.")
41
Regulación de la actividad enzimática
Mecanismos alostéricos Modificaciones covalentes

42
Mecanismos alostéricos
Hay enzimas que pueden adoptar 2 conformaciones interconvertibles llamadas R (relajada) y T (tensa). R es la forma más activa porque se une al sustrato con más afinidad. Las formas R y T se encuentran en equilibrio R <==> T.
 y T (tensa). R es la forma más activa porque se une al sustrato con más afinidad. Las formas R y T se encuentran en equilibrio R <==> T.")
43
Ciertas sustancias tienden a estabilizar la forma R
Ciertas sustancias tienden a estabilizar la forma R. Son los llamados moduladores positivos. El propio sustrato es a menudo un modulador positivo. Las moléculas que favorecen la forma R pero que actúan sobre una región del enzima distinta del centro activo son los activadores alostéricos

44
Las sustancias que favorecen la forma T y disminuyen la actividad enzimática son los moduladores negativos. Si estos moduladores actúan en lugares distintos del centro activo del enzima se llaman inhibidores alostéricos.

45
Modificaciones covalentes
Otras enzimas pasan de una forma menos activa a otra más activa uniéndose covalentemente a un grupo químico de pequeño tamaño como el Pi o el AMP. También se da el caso inverso, en el que un enzima muy activo se desactiva al liberar algún grupo químico. En las enzimas de las vías degradativas del metabolismo, la forma fosforilada es más activa que la no fosforilada, mientras que en las vías biosintéticas ocurre lo contrario.

46
Enzima no fosforilada (Inactiva) Enzima fosforilada (Activa)
 Enzima fosforilada (Activa)")
47
ACTIVACIÓN DE ZIMÓGENOS
Algunas enzimas no se sintetizan como tales, sino como proteínas precursoras sin actividad enzimática. Estas proteínas se llaman proenzimas o zimógenos. Para activarse, los zimógenos sufren un ataque hidrolítico que origina la liberación de uno o varios péptidos. El resto de la molécula protéica adopta la conformación y las propiedades de la enzima activa.

48
Muchas enzimas digestivas se secretan en forma de zimógenos y en el tubo digestivo se convierten en la forma activa. Es el caso de la -quimotripsina, que se sintetiza en forma de quimotripsinógeno

49
Si estas enzimas se sintetizaran directamente en forma activa destruirían la propia célula que las produce. Así, la tripsina pancreática (una proteasa) se sintetiza como tripsinógeno (inactivo). Si por alguna razón se activa en el propio páncreas, la glándula sufre un proceso de autodestrucción (pancreatitis aguda), a menudo mortal.
 se sintetiza como tripsinógeno (inactivo). Si por alguna razón se activa en el propio páncreas, la glándula sufre un proceso de autodestrucción (pancreatitis aguda), a menudo mortal.")
50
Proteínas de la coagulación

51
Coenzimas Son compuestos orgánicos requeridos por las enzimas para realizar su actividad catalítica. Actúan como reactivos de transferencia de grupos. Son específicas para los grupos químicos, a los que se les llama grupos metabólicos móviles, que ellas aceptan y donan. Los grupos metabólicos móviles son fijados al centro reactivo de la coenzima.

53
Algunas coenzimas se sintetizan a partir de metabolitos comunes y nos referimos a ellas como coenzimas metabolitos. El ATP es la más familiar y la más abundante de estas coenzimas.

54
Algunas vitaminas son necesarias para la actuación de determinados enzimas, ya que funcionan como coenzimas que intervienen en distintas rutas metabslicas y , por ello, una deficiencia en una vitamina puede originar importantes defectos metabólicos. En los animales, muchas coenzimas se derivan de las vitaminas B.

55
Enfermedades carenciales C (ácido ascsrbico)
VITAMINAS FUNCIONES Enfermedades carenciales C (ácido ascsrbico) Coenzima de algunas peptidasas. Interviene en la síntesis de colágeno Escorbuto B1 (tiamina) Coenzima de las descarboxilasas y de las enzimas que transfieren grupos aldehidos Beriberi B2 (riboflavina) Constituyente de los coenzimas FAD y FMN Dermatitis y lesiones en las mucosas B3 (ácido pantotinico) Constituyente de la CoA Fatiga y trastornos del sueño B5 (niacina) Constituyente de las coenzimas NAD y NADP Pelagra B6 (piridoxina) Interviene en las reacciones de transferencia de grupos amino. Depresión, anemia B12 (cobalamina) Coenzima en la transferencia de grupos metilo. Anemia perniciosa Biotina Coenzima de las enzimas que transfieren grupos carboxilo, en metabolismo de aminoácidos. Fatiga, dermatitis...
 Coenzima de algunas peptidasas. Interviene en la síntesis de colágeno. Escorbuto. B1 (tiamina) Coenzima de las descarboxilasas y de las enzimas que transfieren grupos aldehidos. Beriberi. B2 (riboflavina) Constituyente de los coenzimas FAD y FMN. Dermatitis y lesiones en las mucosas. B3 (ácido pantotinico) Constituyente de la CoA. Fatiga y trastornos del sueño. B5 (niacina) Constituyente de las coenzimas NAD y NADP. Pelagra. B6. (piridoxina) Interviene en las reacciones de transferencia de grupos amino. Depresión, anemia. B12 (cobalamina) Coenzima en la transferencia de grupos metilo. Anemia perniciosa. Biotina. Coenzima de las enzimas que transfieren grupos carboxilo, en metabolismo de aminoácidos. Fatiga, dermatitis...")
56
Escorbuto Beriberi Pelagra Anemia Perniciosa

57
Coenzima Q (CoQ) Esta coenzima es sintetizada a partir de farnesil pirofosfato. La CoQ es capaz de aceptar y donar uno o dos electrones porque su forma semiquinona es estable. A su forma oxidada se le conoce como ubiquinona o quinona. A su forma reducida se le conoce como QH2, ubiquinol o hidroquinona.

58
Biotina La biotina consiste de un anillo imidazol que está unido en forma cis a la cadena lateral tetrahidrotiofeno del valerato. La enzima holocarboxilasa sintetasa (HCS) cataliza la activación, mediante biotinilación, de cinco carboxilasas en células humanas.
 cataliza la activación, mediante biotinilación, de cinco carboxilasas en células humanas.")
59
NAD y NADP Derivados de la niacina.
Fueron las primeras coenzimas que se reconocieron. Participan en gran cantidad de reacciones metabólicas.

Presentaciones similares