Descargar la presentación
La descarga está en progreso. Por favor, espere

2
Tipos De intrones GU-AG AU-AC Grupo I Grupo II Grupo III
Intrones gemelos pre-tRNA Arqueales pre-mRNA nuclear eucariote pre-rRNA nuclear eucariote RNAs de organelos pre-tRNA nuclear eucariote varios RNAs

3
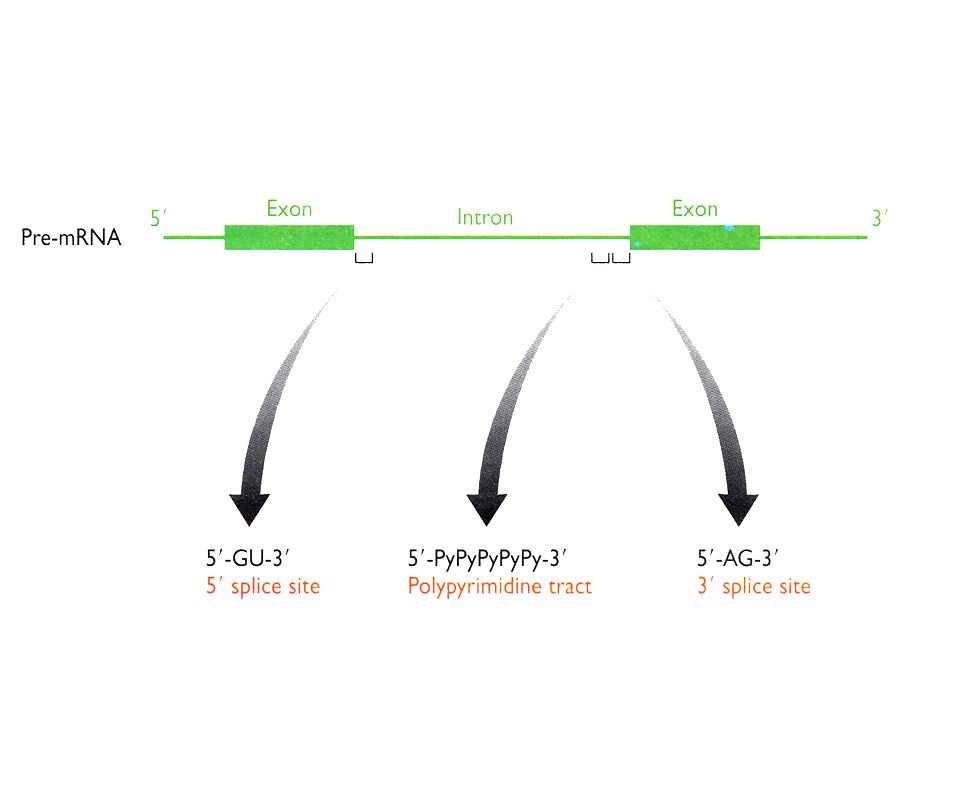
Problema: Definición de los sitios de splicing
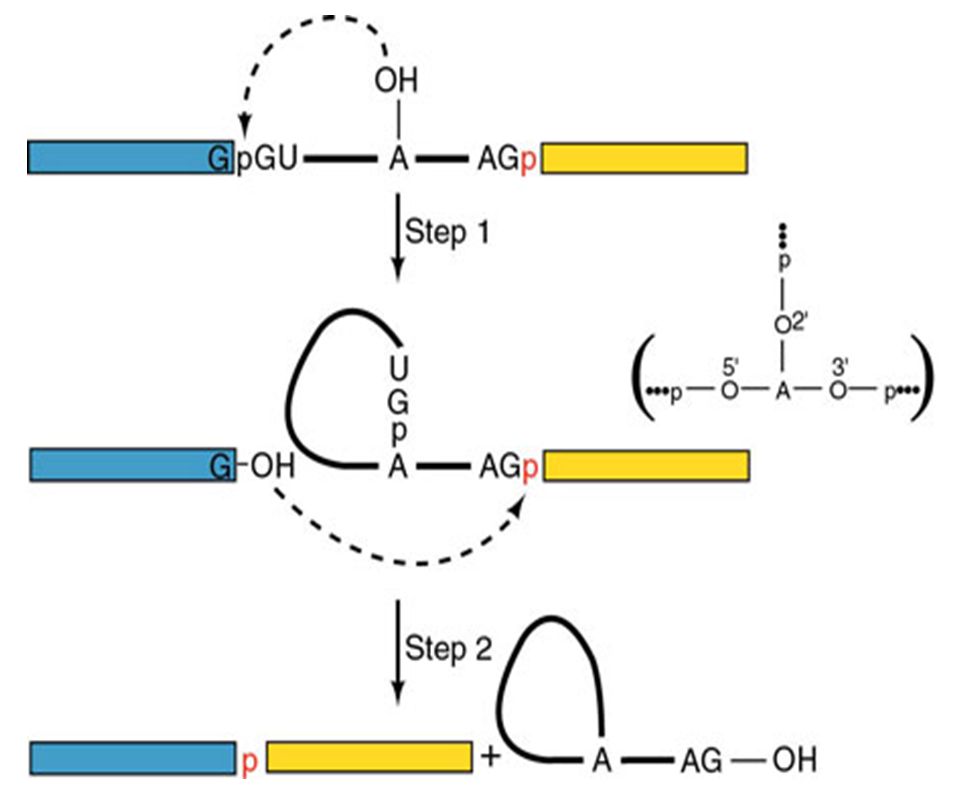
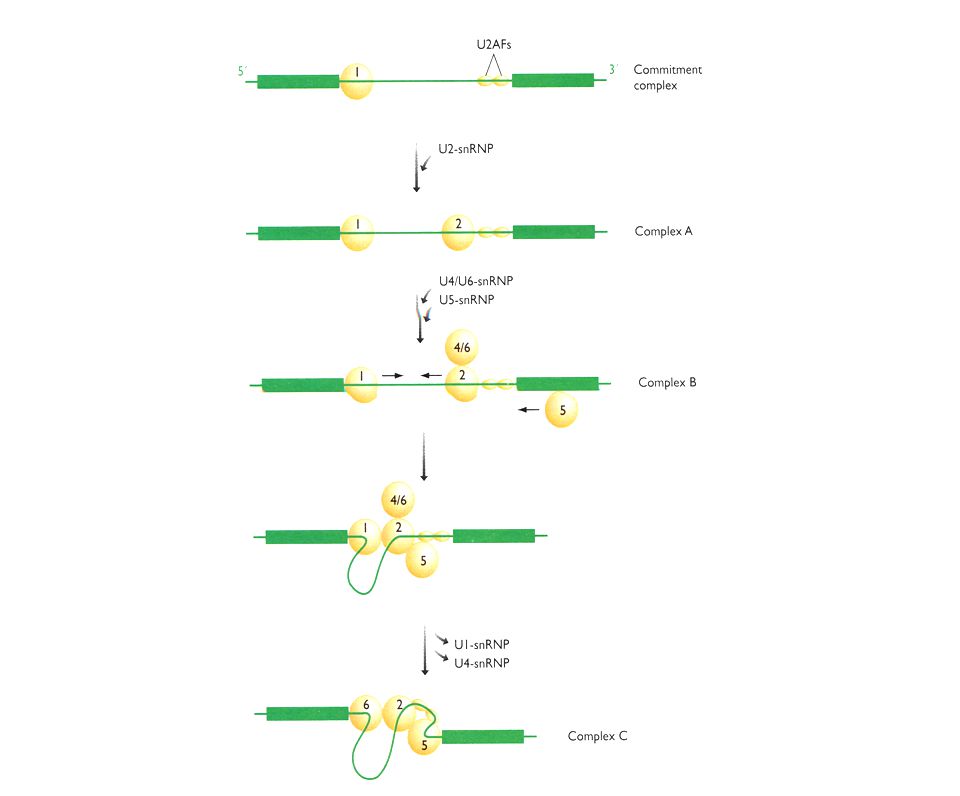
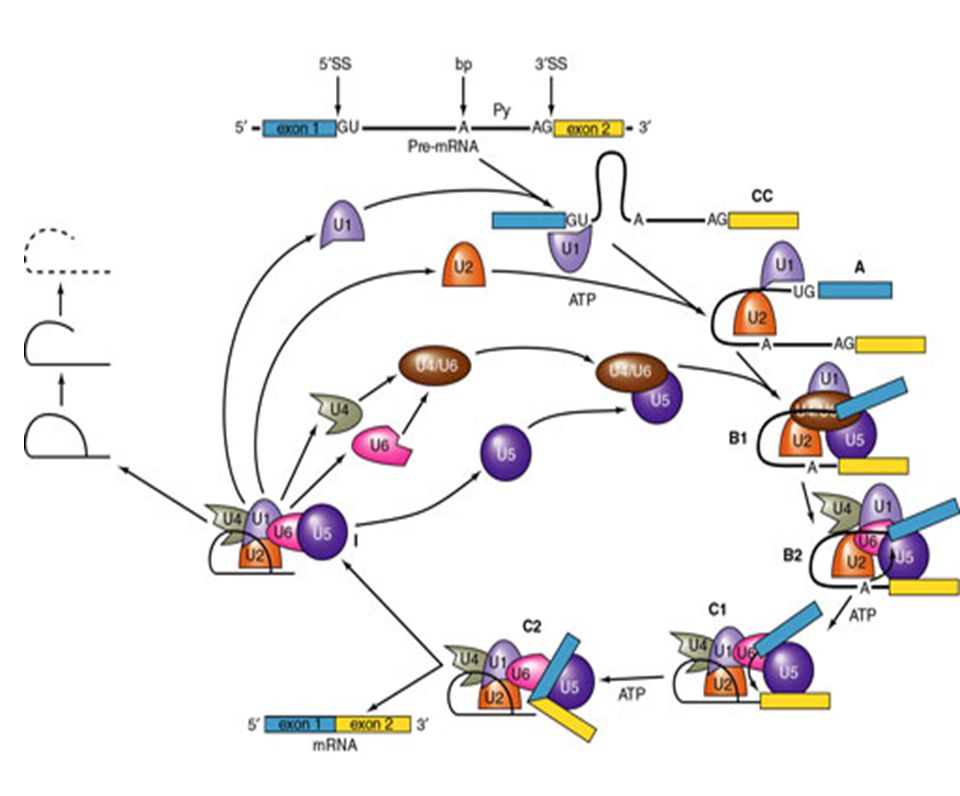
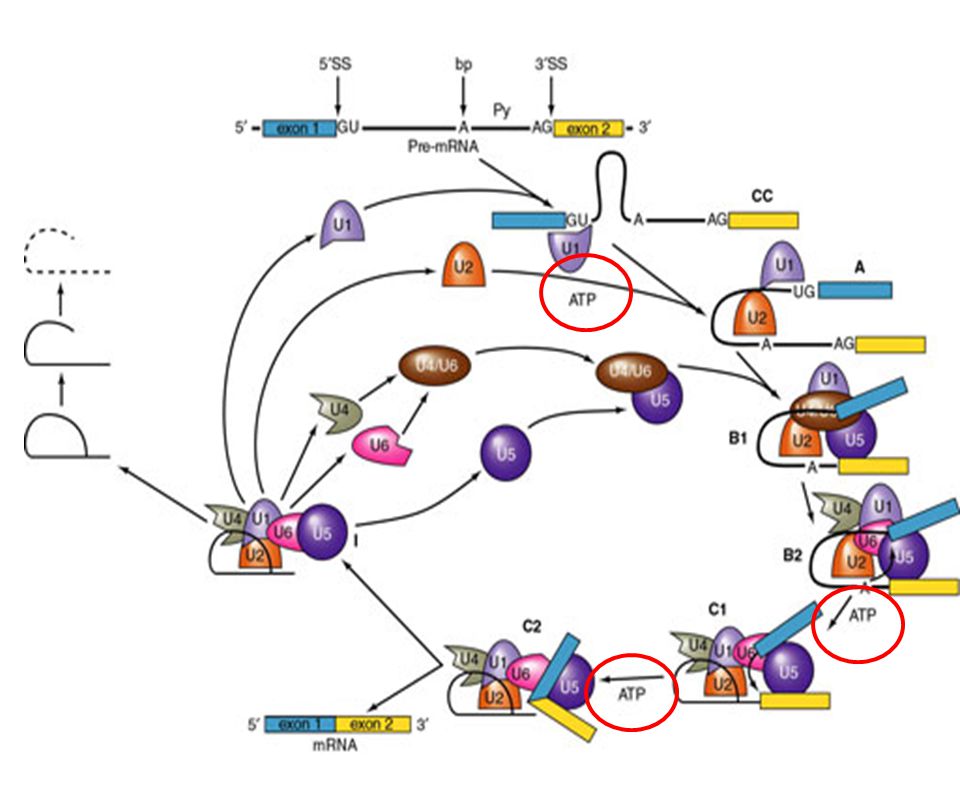
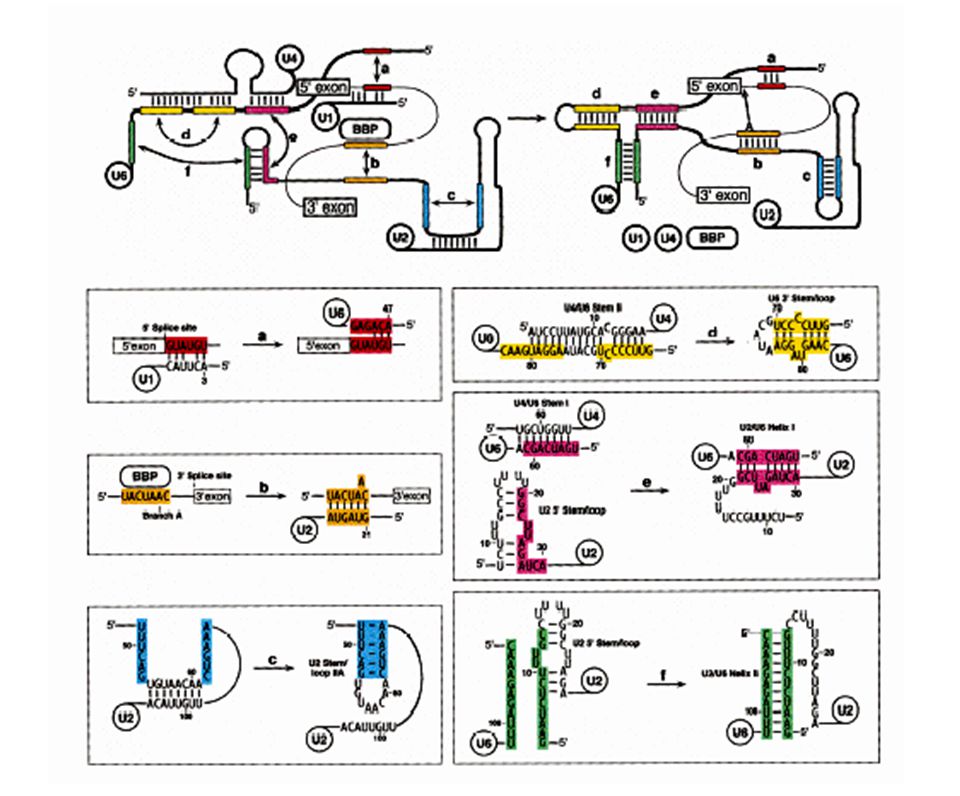
Solución: Secuencias consenso del mRNA en la fronteras entre exón/intrón. Acción de snRNA mediante el apareamiento Watson Crick a sitios cruciales del mRNA. Complejos RNA-proteína (snRNP´s). Ocurrencia: Co-transcripcional. Pol II CTD recluta los factores de splicing. Factores: 5 snRNA: U1 (165 nts), U2 (185), U4 (145), U5 (106), U6 (116). 100 proteínas (organizadas como U1 snRNP, U2 snRNP, etc.). Abundancia – 1 x 106 Catálisis: Transesterificación del RNA
. Ocurrencia: Co-transcripcional. Pol II CTD recluta los factores de splicing. Factores: 5 snRNA: U1 (165 nts), U2 (185), U4 (145), U5 (106), U6 (116). 100 proteínas (organizadas como U1 snRNP, U2 snRNP, etc.). Abundancia – 1 x 106. Catálisis: Transesterificación del RNA.")
7
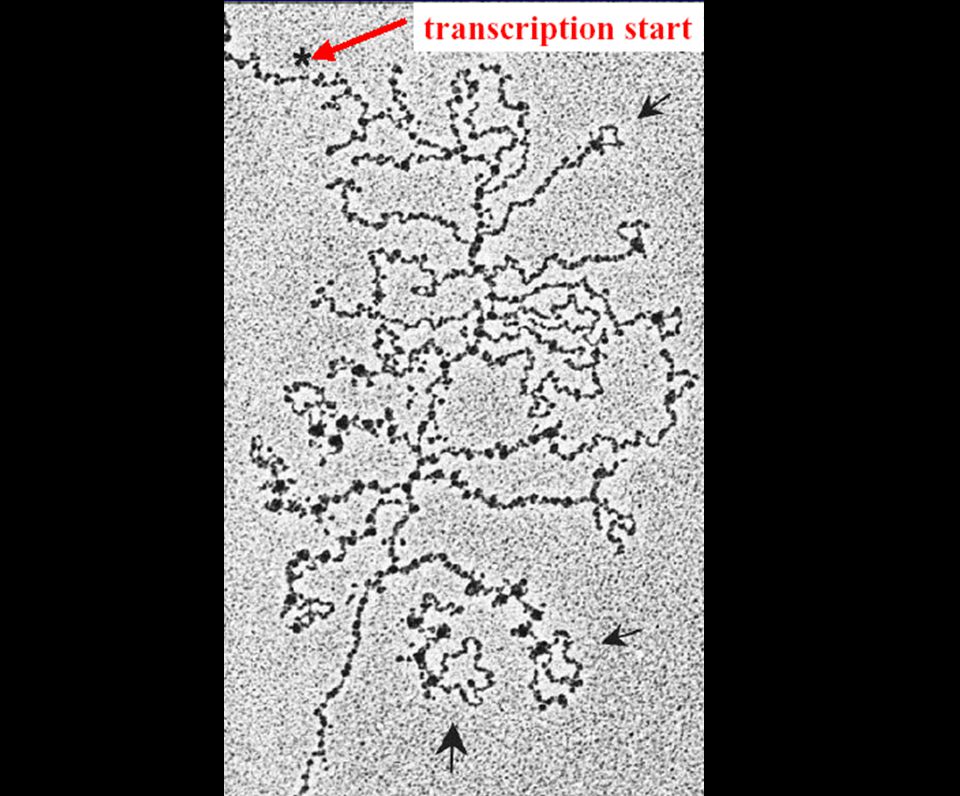
Tres problemas topológicos
¿Cómo identificar sitios de splicing distantes varias kb entre ellos? ¿Cómo identificar los sitios de splicing correctos sin saltarse alguno? ¿Cómo evitar la utilización de sitios crípticos ó espurios?

14
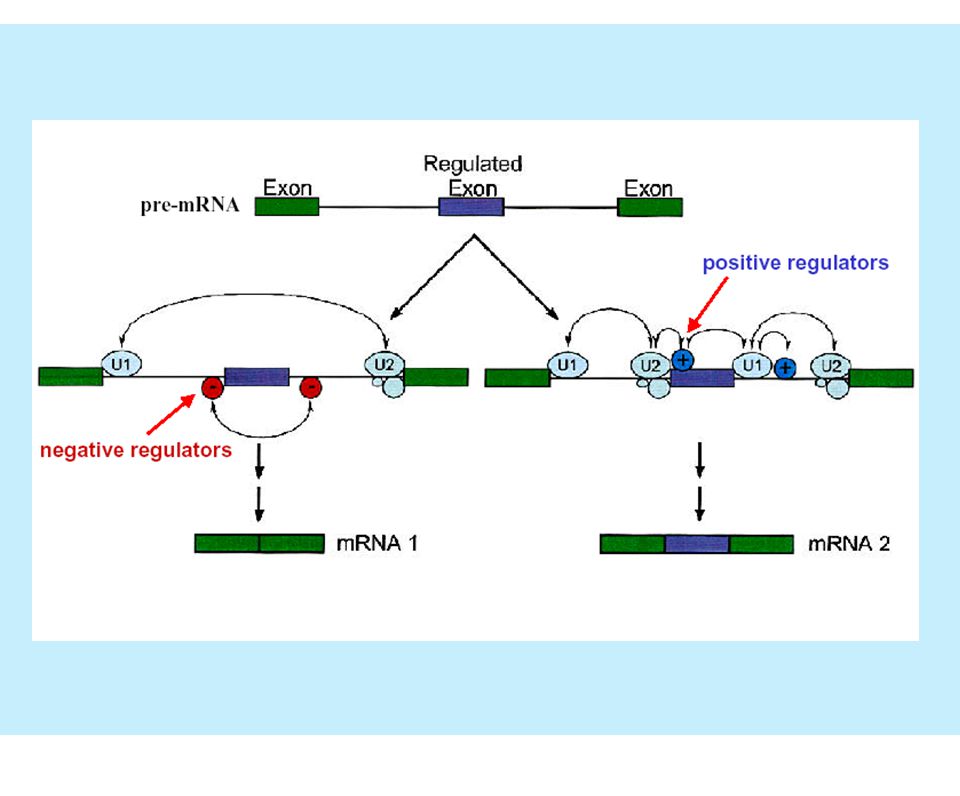
¿Cuáles son los puntos de regulación del splicing?

15
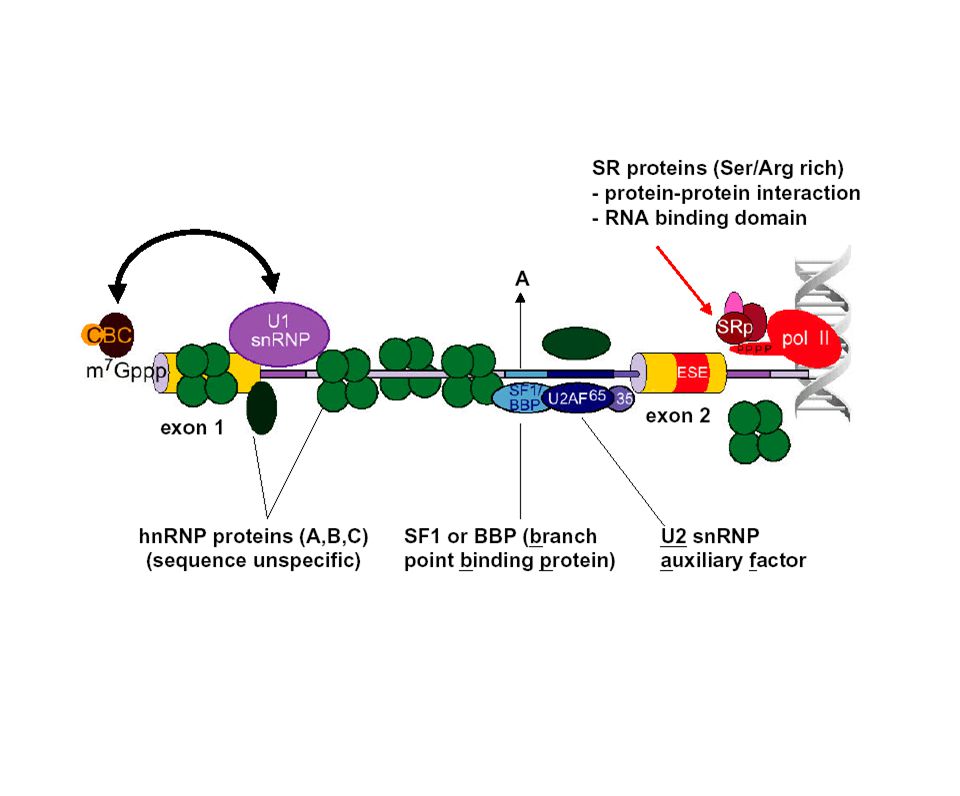
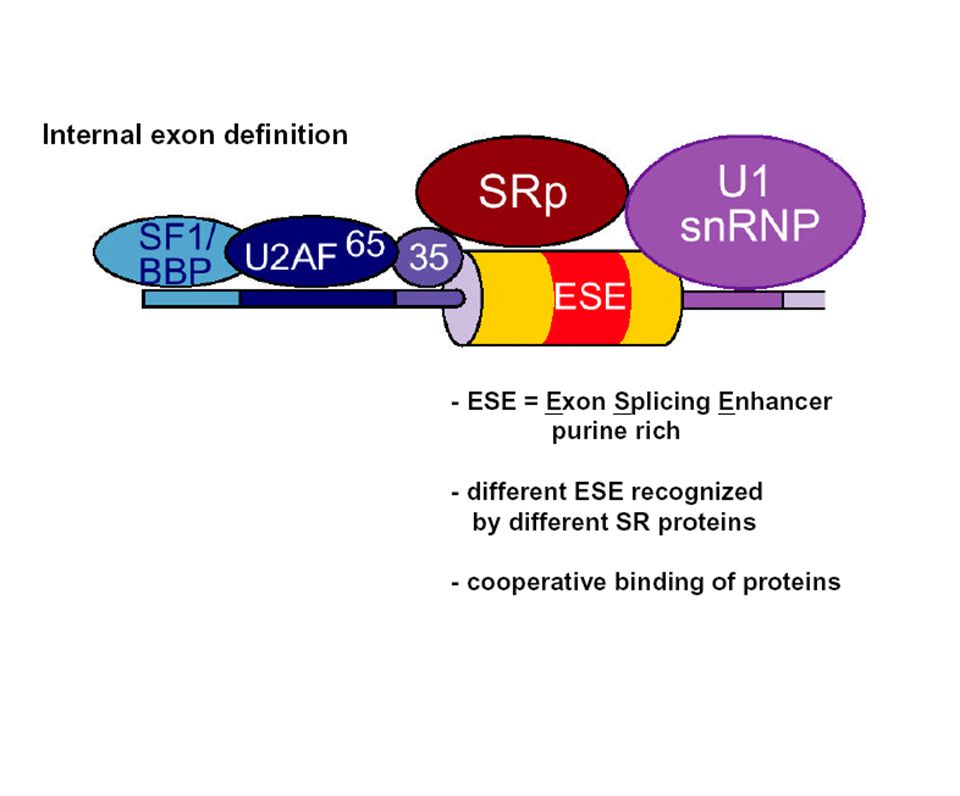
Complejo de definición del exón

17
¿Se remueven los intrones en algún orden específico?

19
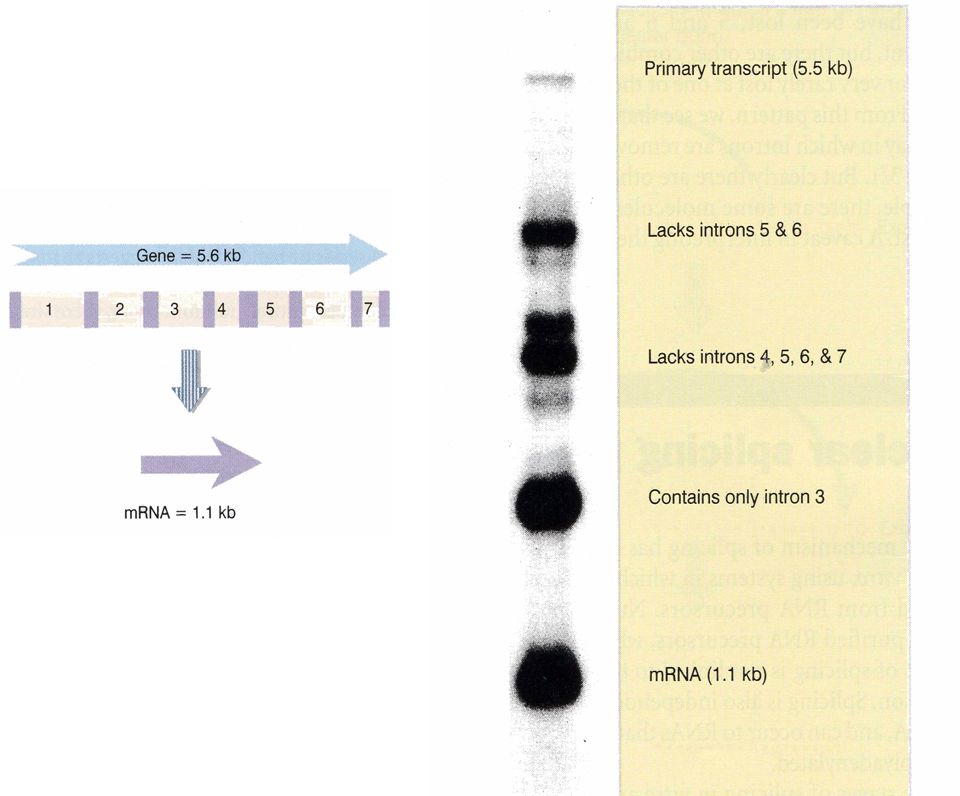
Procesamiento de un exón extra-largo

21
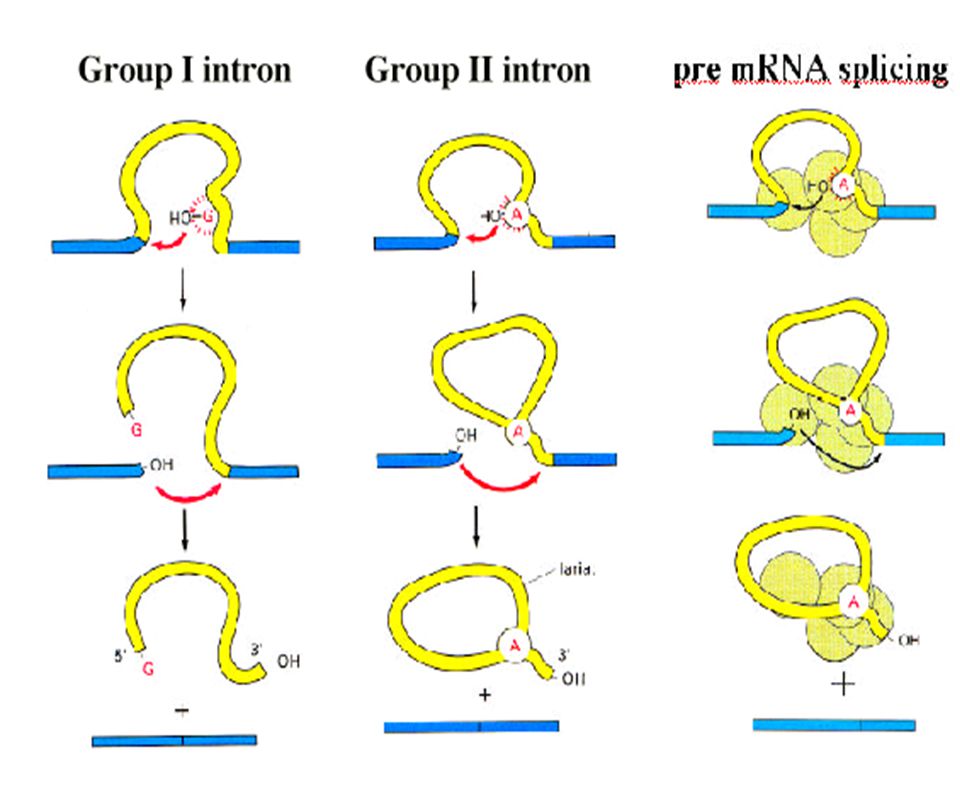
Resumen: procesamiento de intrones de los grupos I y II
Intron del grupo I Intron del grupo II

22
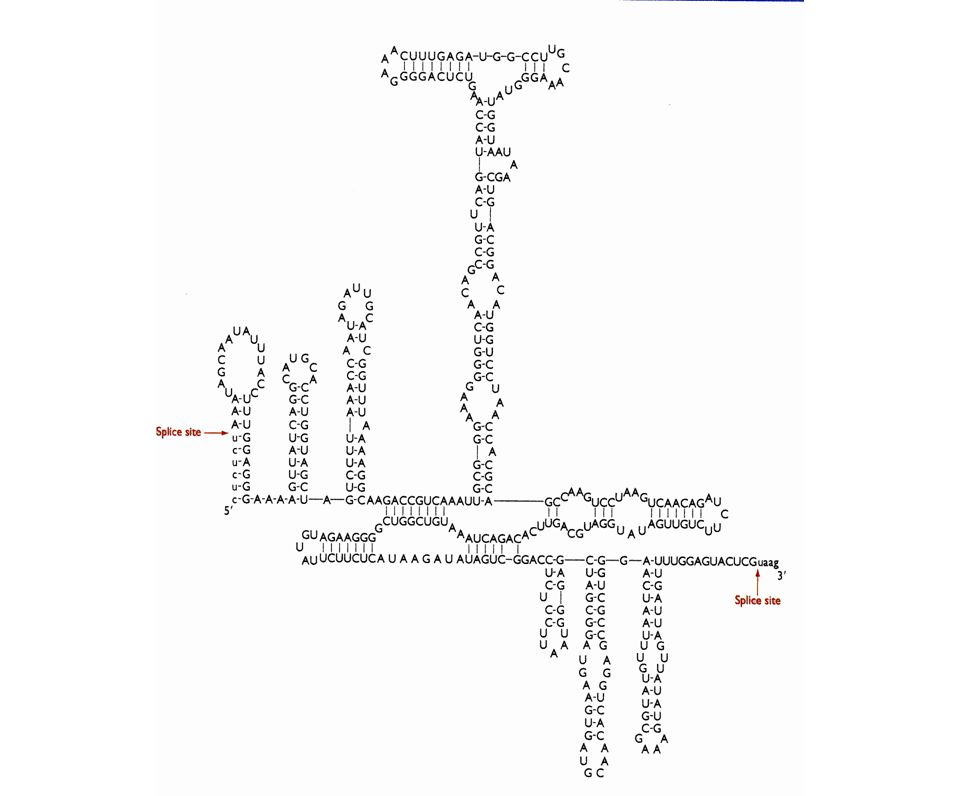
¿Cómo Se Especifican Los Sitios Del Autoprocesamiento?

24
intrón del rRNA de Tetrahymena

25
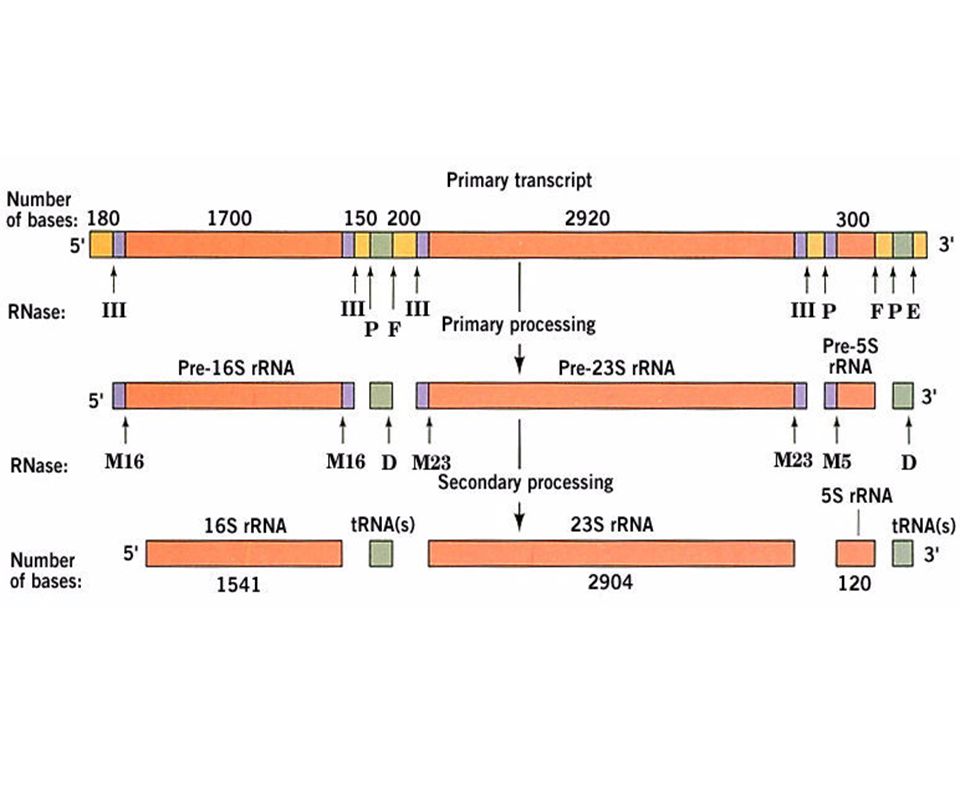
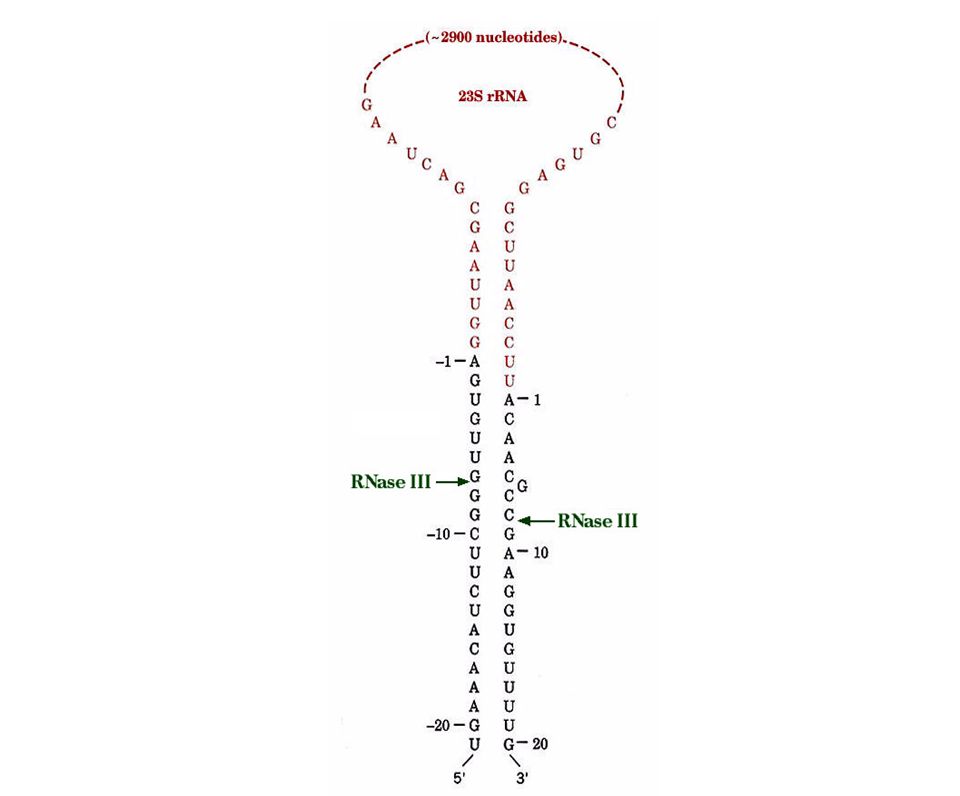
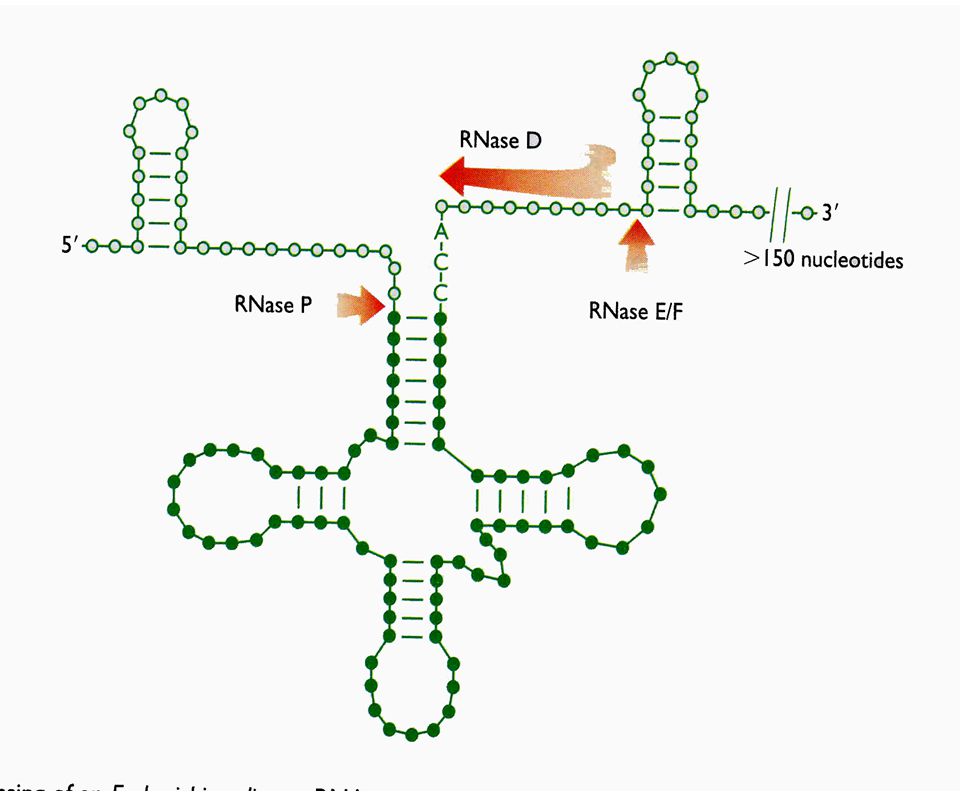
Procesamiento por corte especifico - RNAs no codificantes

29
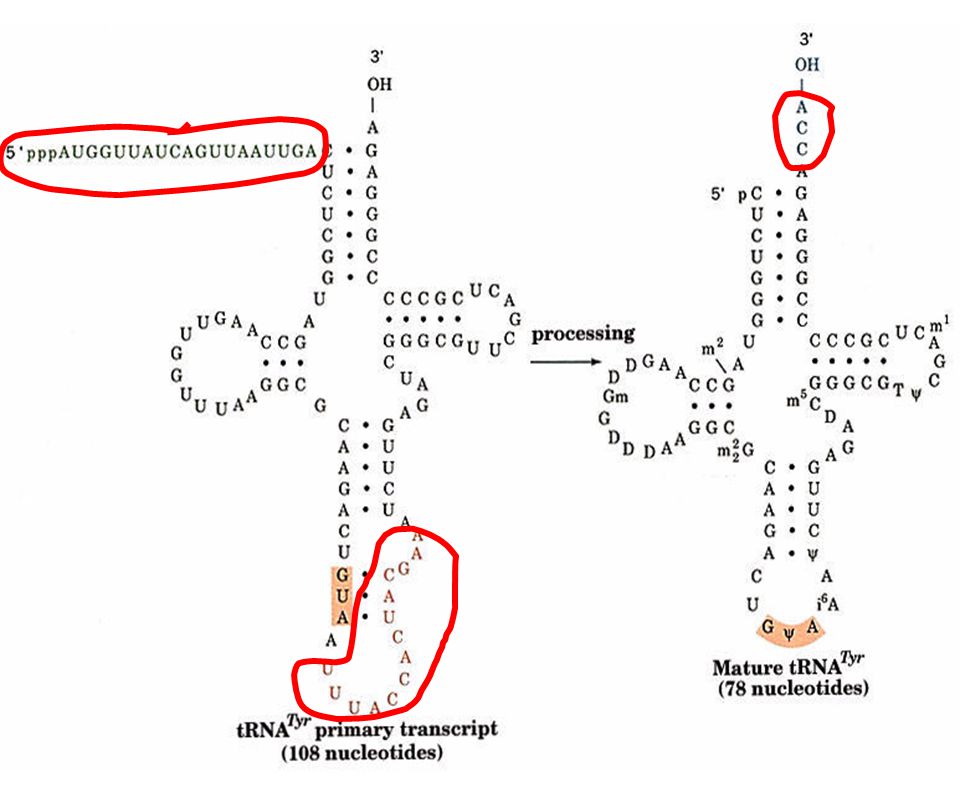
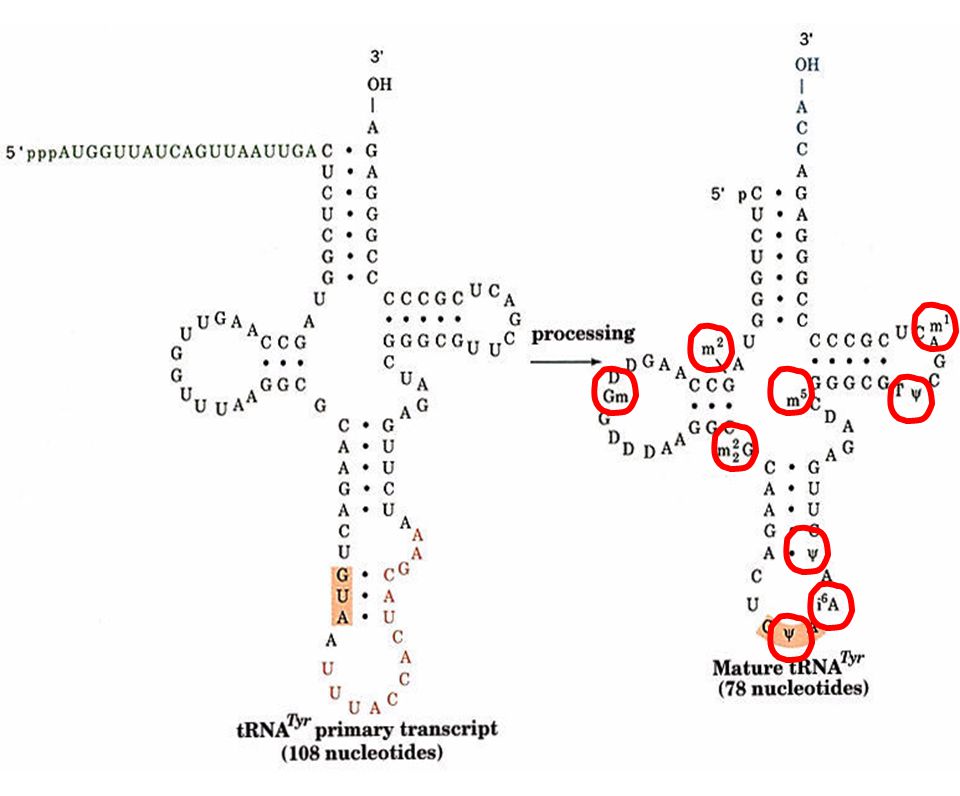
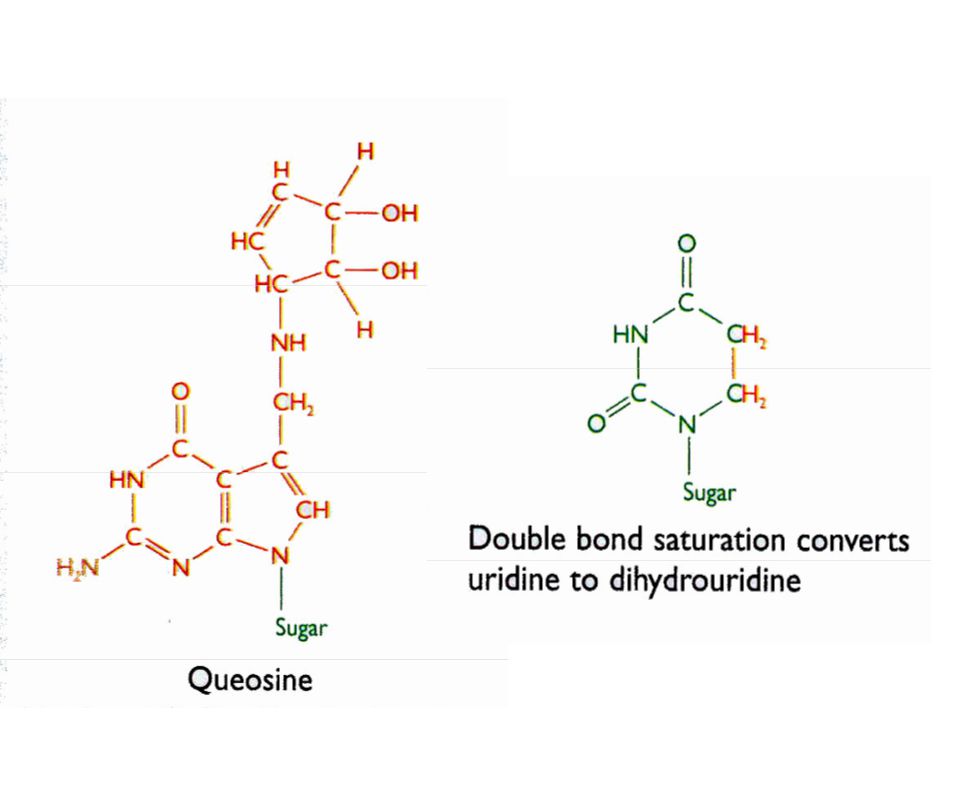
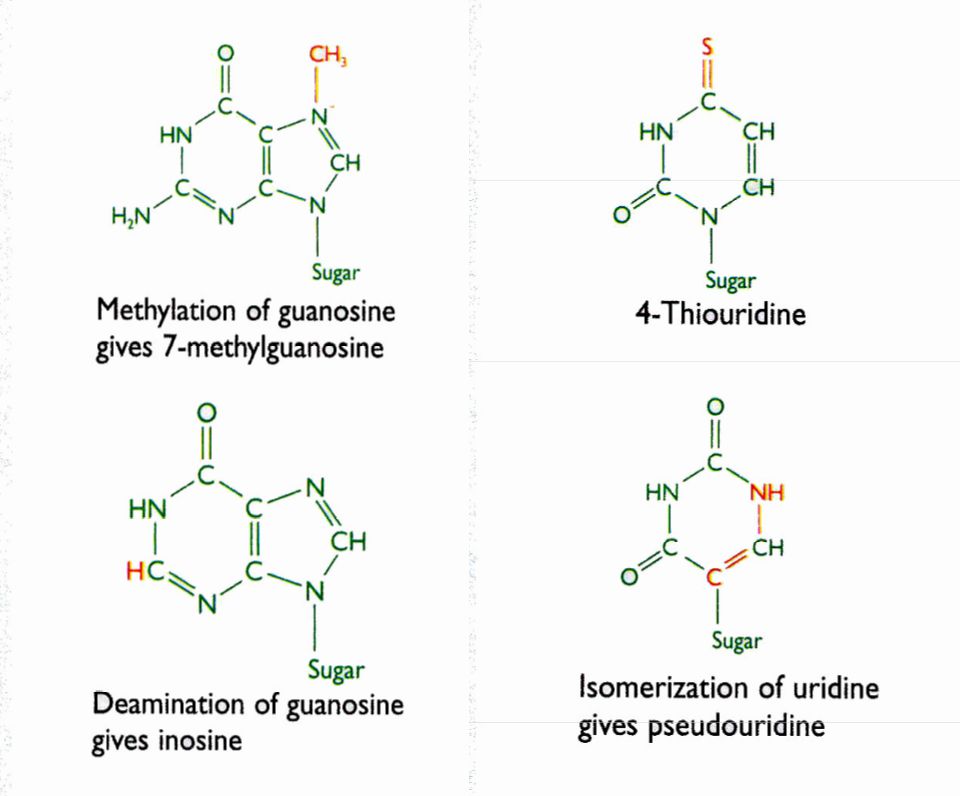
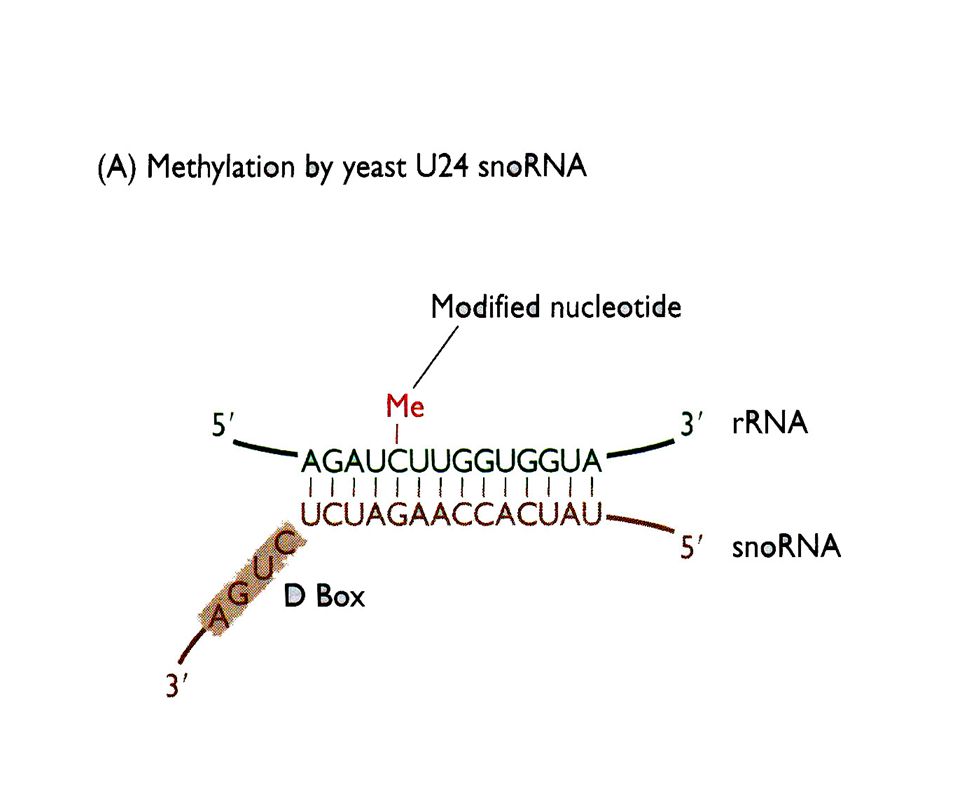
Procesamiento por modificación química de pre r-RNAs y pre t-RNAs

34
Un pre rRNA humano sufre 106 metilaciones y 95 pseudouridilaciones de manera específica y con un patrón extremadamente conservado

37
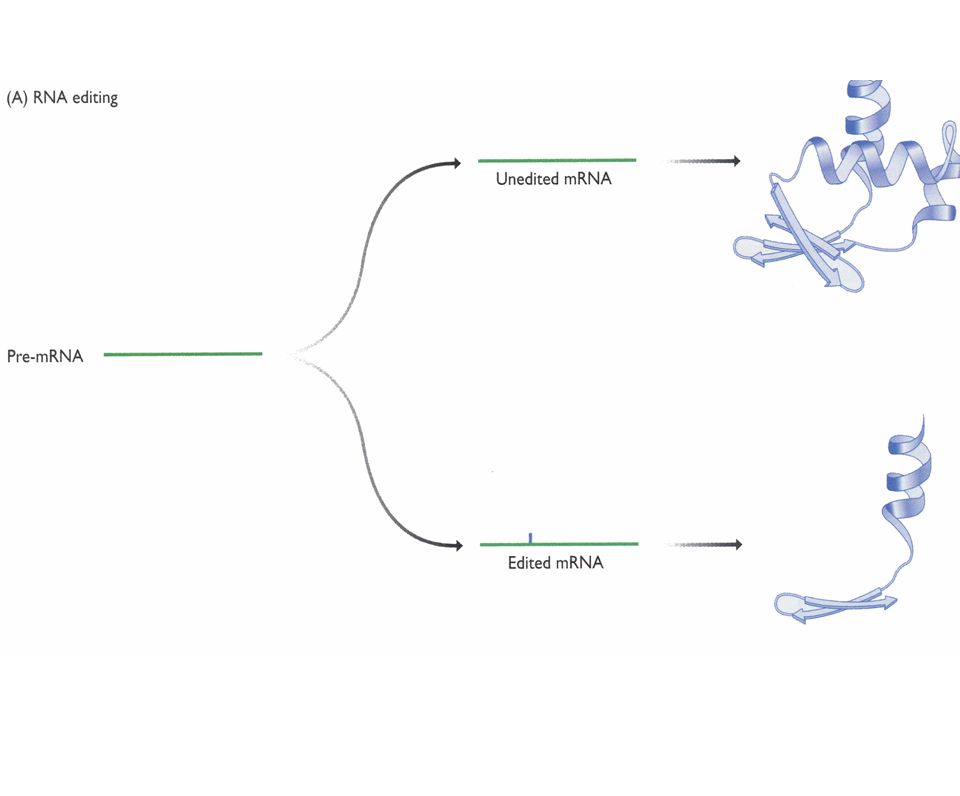
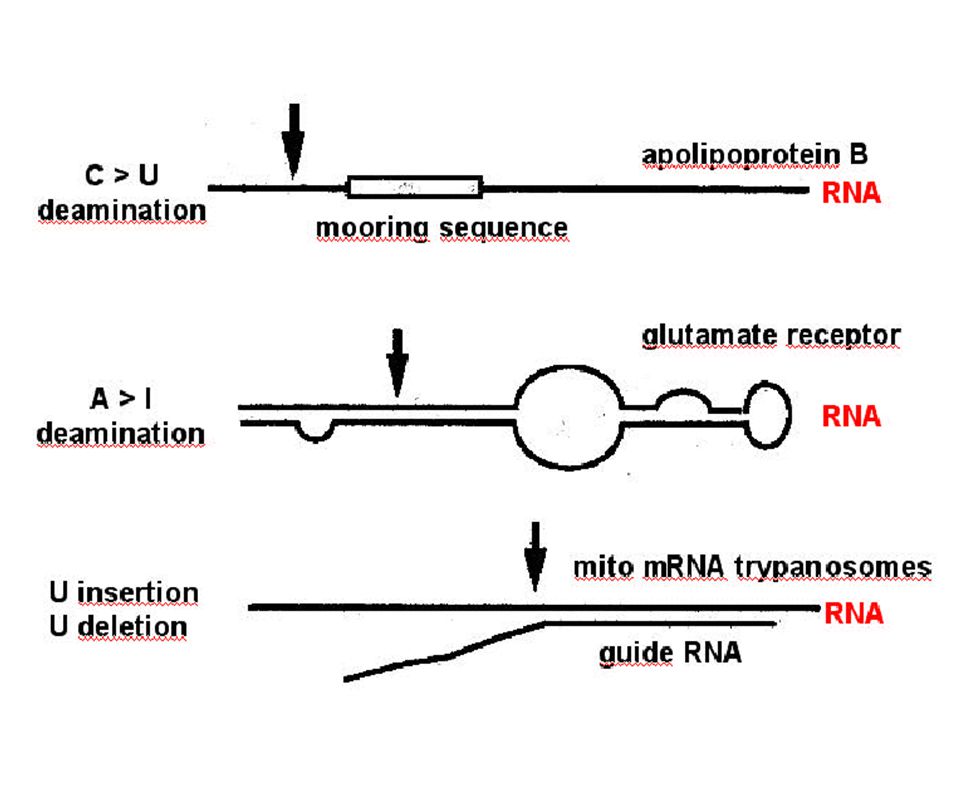
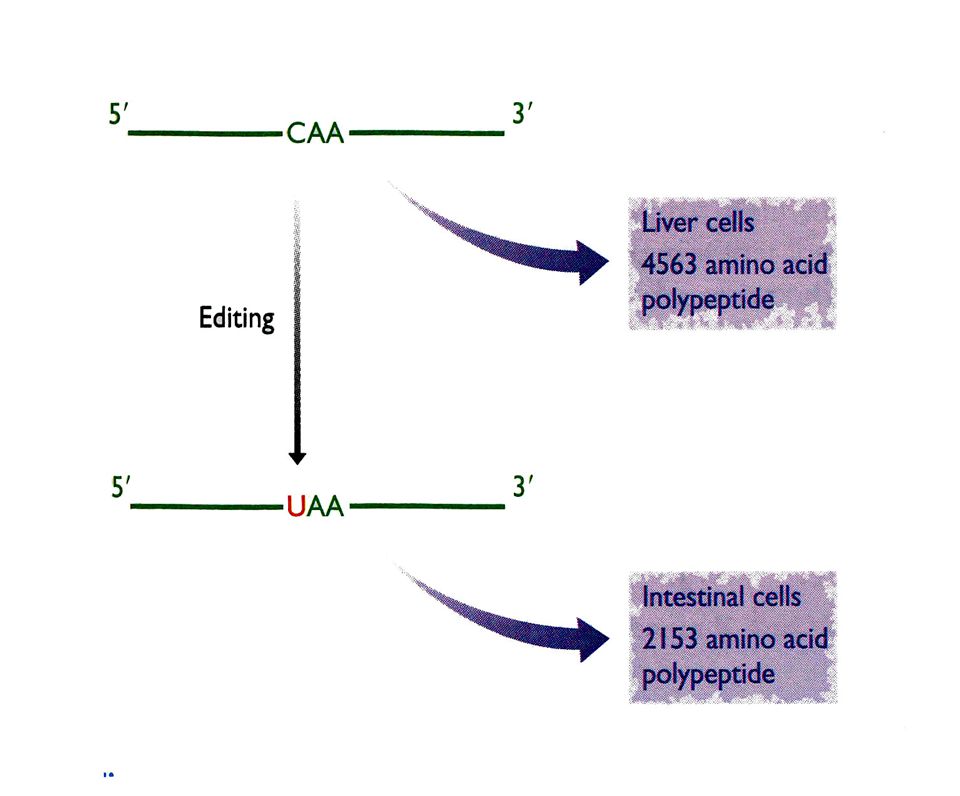
Edicion de RNAs

42
¿Que sucede cuando hay intrones múltiples en un gene?

43
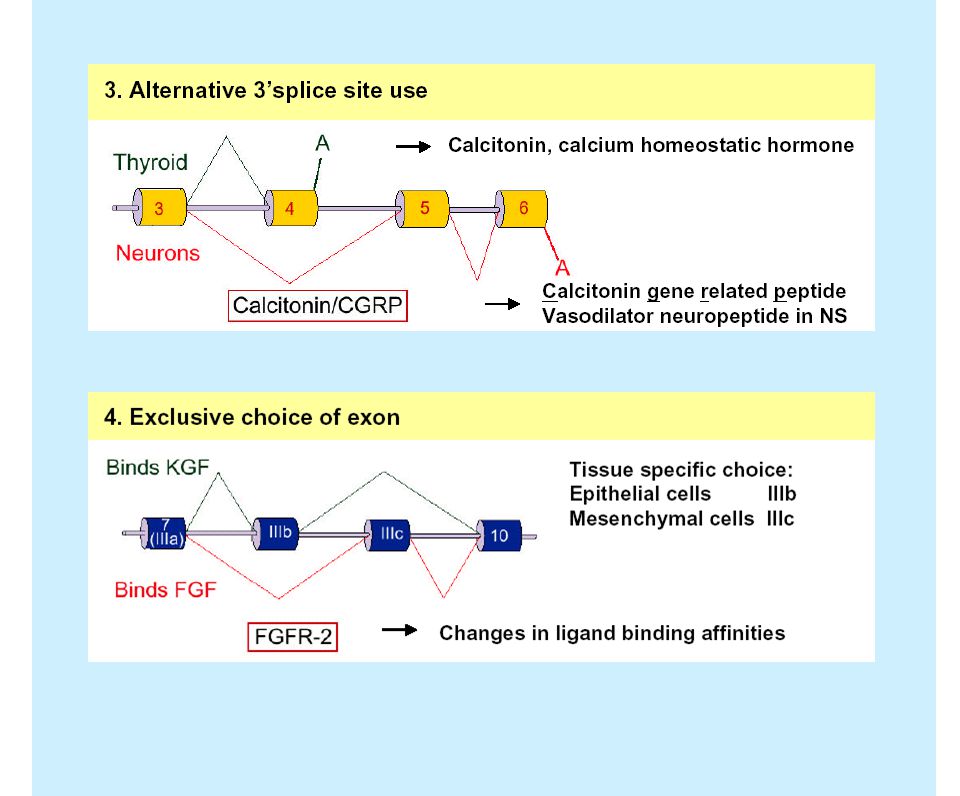
Ejemplos de splicing alternativo

47
Transporte del mRNA procesado
Factores de especificidad para el procesamiento y la poliadenilación

48
Subunidades ribosomales 5/min
mRNA /min Histonas /min Otras proteínas /min Proteínas ribosomales 150/min

49
FIG. 1. A, diagram of the nuclear pore complex (center) and the proteins that participate in nucleocytoplasmic transport in S. cerevisiae. Nups are the structural components of the NPC; these include the FG Nups, the non-FG Nups (which lack FG peptide repeats), and the nuclear pore membrane proteins (POMs). Kaps bind and translocate cargo across the NPC; these include the importins (Kaps dedicated to nuclear import), the exportins (Kaps dedicated to nuclear export), and the transportins (Kaps that carry cargo in both directions across the NPC). The Gsp1p GTPase (Ran in vertebrates) and its effectors (Rna1p is a GTPase-activating protein; Prp20p and Mog1p are guanine nucleotide exchange factors; Yrb1p is a guanine nucleotide dissociation inhibitor) impart directionality to the transport process by modulating the interaction between Kaps and Nups. B, flow chart of our proteomic strategy for deciphering and characterizing networks of protein interactions at the nuclear pore complex. Cells are grown under any of a variety of conditions and are lysed. Soluble proteins are separated from insoluble proteins by differential centrifugation. Proteins in the soluble pool are mixed with Sepharose beads coated with a GST fusion (GST-nucleoporin, GST-karyopherin, etc.). After washing beads, specifically bound proteins are eluted with salt and resolved by SDS-PAGE. Individual proteins are then identified by mass spectrometric analysis. Identified proteins are produced in recombinant form and tested for their ability to bind directly to the GST fusion in the absence of other proteins or in the presence of adapter proteins. Direct interactions detected in vitro are then confirmed in vivo using yeast two-hybrid assay. Deciphering networks of protein interactions at the nuclear pore complex. Allen et al. Molecular and Cellular Proteomics (2002) 1.12 pp
 1.12 pp")
50
FIG. 8. Summary of protein interactions identified thus far using the proteomic strategy described here. The diagram shows vertical lists of proteins captured by immobilized Nups and Kaps (shown atop in bold). Nups are clustered according to their location in the NPC. Solid line boxes denote protein complexes that have been confirmed; dotted line boxes indicate likely protein complexes based on the published literature. Proteins listed below the horizontal lines in each row have not yet been studied in detail. a, Ref. 4; b, Ref. 5; d, N. P. C. Allen and M. Rexach, unpublished data; e, Ref. 10; f, Ref. 8. Deciphering networks of protein interactions at the nuclear pore complex. Allen et al. Molecular and Cellular Proteomics (2002) 1.12 pp
 1.12 pp")
51
Computer generated diagrams depicting hypothetical roles for the interactions characterized here. A. The Nup84p complex (in yellow) may interact with various FG Nups in the central FG Nup meshwork (in light blue) to help mantain a seal between the scaffold rings and the filamentous FG Nups. Deciphering networks of protein interactions at the nuclear pore complex. Allen et al. Molecular and Cellular Proteomics (2002) 1.12 pp
 1.12 pp")
52
Control de la estabilidad del RNA y su degradación

53
Señales que identifican al mRNA procesado
Models for mRNA surveillance. (a) Nuclear assembly and processing generates complex RNPs. Exon junction markers are indicated. (b) Upon exportat from the nucleos, eIF4G binds to thge CBC and translation is initiated mRNA turnover. Mitchell and Tollervey (2001) Current Opinion in Cell Biology 13: (b)
 Nuclear assembly and processing generates complex RNPs. Exon junction markers are indicated. (b) Upon exportat from the nucleos, eIF4G binds to thge CBC and translation is initiated. mRNA turnover. Mitchell and Tollervey (2001) Current Opinion in Cell Biology 13: (b)")
54
Remoción de señales durante la primera ronda de traducción de un mensajero silvestre
During elongation, marker proteins located within the ORF are dissociated. Upon translation termination, Upf1p is recrited by the release factors (RF) and a surveillance complex (SC) that includes Upf1p then translocates trhough the 3’ UTR mRNA turnover. Mitchell and Tollervey (2001) Current Opinion in Cell Biology 13:
 and a surveillance complex (SC) that includes Upf1p then translocates trhough the 3’ UTR. mRNA turnover. Mitchell and Tollervey (2001) Current Opinion in Cell Biology 13:")
55
Circularización del mRNA después de la primera ronda de traducción
If only correct marker proteins are encountered the mature mRNP structure is formed. eIF4E replaces CBC and the interaction between PABP and eIF4G circularizes the mRNA mRNA turnover. Mitchell and Tollervey (2001) Current Opinion in Cell Biology 13:
 Current Opinion in Cell Biology 13:")
56
Activación de los factores de liberación por codones de terminación prematuros
Interaction of the surveillance complexes with inappropriate proteins triggers decaspping and rapid degrdation. Upf2 and 3p may assemble with Upf1p before scanning or only after scanning is inhibited at the mark. mRNA turnover. Mitchell and Tollervey (2001) Current Opinion in Cell Biology 13:
 Current Opinion in Cell Biology 13:")
57
La formación de complejos de RF, SC y marcadores del sitio de procesamiento activan a la enzima que remueve el cap mRNA turnover. Mitchell and Tollervey (2001) Current Opinion in Cell Biology 13:
 Current Opinion in Cell Biology 13:")
Presentaciones similares













>")








