Descargar la presentación
La descarga está en progreso. Por favor, espere

1
LAS NEURONAS

2
Las neuronas: Son las más características y más estudiadas por la relación de sus propiedades con las funciones del sistema nervioso. Existen en enorme número * 106, 100 billones. Funcionalmente polarizadas. Esto es, reciben información por uno de sus extremos, dendrítico y la entregan por otro, extremo axónico. Tienen una enorme capacidad de comunicarse con otras células, especialmente con otras neuronas. Una neurona está compuesta por: Las dendritas El cuerpo celular o soma El axón

3
Neurona: el cuerpo celular o soma, las dendritas, que reciben la información desde otras neuronas, el axón, por donde el impulso nervioso viaja hacia otras células. El axón de esta neurona en particular está mielinizado. La mielina está formada por células gliales que envuelven el axón para favorecer la conducción de la señal nerviosa. El axón se ramifica hacia terminales o botones sinápticos.

4
Las dendritas y el axón constituyen los procesos neuronales.
Las dendritas nacen del soma o cuerpo neuronal y pueden ser muy abundantes y ramificadas. Son las que reciben la información. El axón nace del soma, en la región del montículo axónico, que se continúa con el segmento inicial del axón que es donde se generan los potenciales de acción. Un potencial de acción es una señal de electricidad negativa que viaja por el axón a una velocidad variable, según el tipo de axón, hasta alcanzar la región terminal donde induce liberación de una señal o mensaje químico, el neurotransmisor. Los axones pueden ser muy cortos o alcanzar longitudes de más de un metro. En algunas regiones, el axón emite una "colateral" (una ramificación) que va a inervar una neurona vecina (por, ejemplo la interneurona de Renshaw) o vuelve a la región del soma, colateral recurrente.
 que va a inervar una neurona vecina (por, ejemplo la interneurona de Renshaw) o vuelve a la región del soma, colateral recurrente.")
5
Las células gliales: Son veces más numerosas que las neuronas y las rodean. Presentan ramificaciones, a veces muy escasas, y cortas que se unen a un cuerpo pequeño. Aunque no se las considera esenciales para el procesamiento y conducción de la información se les atribuye funciones muy importantes para el trabajo neuronal: Soporte mecánico y aislamiento de las neuronas. Ellas aíslan el axón, sin impedir el proceso de autogeneración del potencial de acción, con lo que se logra acelerar la velocidad de propagación de esta señal. Mantienen la constancia del microambiente neuronal, eliminando exceso de neurotransmisores y/o de sus metabolitos y de iones Guían el desarrollo de las neuronas y parecen cumplir funciones nutritivas para este tipo de células.

6
IMPULSO NERVIOSO

7
Membrana plasmática: recordando su estructura

8
Membrana plasmática: origen de su polaridad

9
Pieza clave: Bomba Sodio - Potasio

10
Potencial de reposo y potencial de acción

11
Pistas para explicar potencial de acción: permeabilidad del Na+ y K+

12
Origen del potencial de reposo

13
Origen del potencial de acción

14
Aceleración del impulso nervioso

15
SINAPSIS

16
Sinapsis: zona especializada de contacto entre las neuronas donde tiene lugar la transmisión de la información. → zona de contacto especializada entre una célula presináptica y una célula postsináptica (nerviosa, muscular o glandular), siendo el flujo de información de la 1ª a la 2ª. → Tipos: Eléctricas Químicas
, siendo el flujo de información de la 1ª a la 2ª. → Tipos: Eléctricas. Químicas.")
17
Sinapsis eléctricas El potencial de acción se transmite a la neurona postsináptica por el flujo directo de corriente: continuidad entre citoplasmas. La distancia entre membranas es de unos 3 nm. El flujo de corriente pasa a través de uniones comunicantes (gap junctions formadas por conexinas. Es bidireccional. El hexámero de conexinas forma el conexón. Función: desencadenar respuestas muy rápidas. Sinapsis eléctricas Representan una pequeña fracción de la totalidad de las sinapsis existentes. Para que tengan lugar debe existir una continuidad entre los citoplasmas celulares. Esto es posible mediante la unión en forma de gap-junctions o uniones en hendidura o unione comuinicantes, las cuales dan lugar a la formación de un pequeño poro que permite el paso de iones. Este poro está formado por 6 proteínas, denominadas conexinas, cuyo agrupamiento es conocido como conexón. El paso de iones a través del conexón es bidireccional por lo que la transmisión de información se produce en ambos sentidos. Los canales de conexones no se encuentran permanentemente abiertos, se abren y se cierran, de hecho la entreda de H+ o de Ca2+ así como la despolarización de una o de ambas células facilita la apertura. Aunque estte tipo de sinapsis se encuentra distribuida por los sistemas nerviosos central y periférico de invertebrados y mamíferos este tipo de uniones no son restrictivas del tejido neuronal sino que también han sido observadas por ejemplo en riñón, hígado, estómago páncreas, corazón, células de músculo liso intestinal y células epteliales del cristalino..

18
Sinapsis químicas Liberación de un neurotransmisor (NT) cuando llega el potencial de acción al terminal presináptico El NT difunde por la hendidura sináptica hasta encontrar los receptores postsinápticos Unidireccional Existe retraso sináptico (0,5 ms). Distancia entre membrana pre y postsináptica: nm Sinapsis químicas A diferencia de las sinapsis eléctricas, las químicas son mucho mas abundantes. La transmisión de la información se realiza mediante la liberación, por parte de la neurona presináptica, de un neurotransmisor químico. Esto implica que la transmisión de la información, a diferencia de las eléctricas, es: · Unidireccional · Existe retraso sináptico, ya que desde que se estimula la célula presináptica hasta la detección del efecto en la postsináptica debe de producirse la entrada de Ca2+ para que se estimule la liberación de las vesículas sinápticas, la liberación de neurotransmisor mediante exocitosis y la interacción del mismo con la neurona postsináptica. · La distancia entre las membranas pre- y postsináptica está en el intervalo de 30 a 400nm. Existen diferentes tipos de sinapsis químicas. La mayor parte poseen una serie de características en común: En la terminación nerviosa de la célula presináptica hay vesículas con sustancias neurotransmisoras o neuromoduladoras. Las vesículas que contienen los clásicos neurotransmisores formados por pequeñas moléculas, como la acetilcolina o la norepinefrina, son de reducido tamaño (unos 50nm de diámetro) y se suelen acumular cerca de las áreas especializadas de liberación en la cara interna de la membrana presináptica. A estas zonas electrondensas se les denomina zonas activas. Las vesículas que contienen neuropéptidos son de mayor tamaño y se distribuyen por toda la terminación nerviosa. En muchas terminaciones pueden aparecer conjuntamente ambos tipos vesiculares. El Potencial de acción en la neurona presináptica abre canales Ca2+ dependientes de voltaje concentrados cerca de las zonas activas. El aumento de Ca2+ en el interior celular es el desencadenante de la liberación del neurotransmisor mediante exocitosis en la hendidura sináptica (separación entre las células pre y postsináptica de entre 20 y 40nm de anchura). la zona más próxima a la hendidura presináptica, se acumulas mitocondrias, las cuales generarán la energía necesaria para la exocitosis. La sustancia neurotransmisora difunde a través de la hendidura sináptica hasta unirse a unas proteínas específicas, los receptores de neurotransmisor en la membrana postsináptica. La unión
. Distancia entre membrana pre y postsináptica: nm. Sinapsis químicas. A diferencia de las sinapsis eléctricas, las químicas son mucho mas abundantes. La transmisión de la información se realiza mediante la liberación, por parte de la neurona presináptica, de un neurotransmisor químico. Esto implica que la transmisión de la información, a diferencia de las eléctricas, es: · Unidireccional. · Existe retraso sináptico, ya que desde que se estimula la célula presináptica hasta la detección del efecto en la postsináptica debe de producirse la entrada de Ca2+ para que se estimule la liberación de las vesículas sinápticas, la liberación de neurotransmisor mediante exocitosis y la interacción del mismo con la neurona postsináptica. · La distancia entre las membranas pre- y postsináptica está en el intervalo de 30 a 400nm. Existen diferentes tipos de sinapsis químicas. La mayor parte poseen una serie de características en común: 1- En la terminación nerviosa de la célula presináptica hay vesículas con sustancias neurotransmisoras o neuromoduladoras. Las vesículas que contienen los clásicos neurotransmisores formados por pequeñas moléculas, como la acetilcolina o la norepinefrina, son de reducido tamaño (unos 50nm de diámetro) y se suelen acumular cerca de las áreas especializadas de liberación en la cara interna de la membrana presináptica. A estas zonas electrondensas se les denomina zonas activas. Las vesículas que contienen neuropéptidos son de mayor tamaño y se distribuyen por toda la terminación nerviosa. En muchas terminaciones pueden aparecer conjuntamente ambos tipos vesiculares. 2- El Potencial de acción en la neurona presináptica abre canales Ca2+ dependientes de voltaje concentrados cerca de las zonas activas. El aumento de Ca2+ en el interior celular es el desencadenante de la liberación del neurotransmisor mediante exocitosis en la hendidura sináptica (separación entre las células pre y postsináptica de entre 20 y 40nm de anchura). 3- la zona más próxima a la hendidura presináptica, se acumulas mitocondrias, las cuales generarán la energía necesaria para la exocitosis. 4- La sustancia neurotransmisora difunde a través de la hendidura sináptica hasta unirse a unas proteínas específicas, los receptores de neurotransmisor en la membrana postsináptica. La unión")
19
Organización del botón sináptico
Terminal nervioso Vaina de mielina Citoesqueleto Vesículas sinápticas inmaduras Vesículas sinápticas maduras (aptas para la exocitosis) Vesículas sináptica en exocitosis Neurotransmisor Espacio o hendidura sináptica Membrana presináptica Eudosoma Vesícula sináptica en recuperación Canales de calcio
 Vesículas sináptica en exocitosis. Neurotransmisor. Espacio o hendidura sináptica. Membrana presináptica. Eudosoma. Vesícula sináptica en recuperación. Canales de calcio.")
20
Sinapsis química

21
Interacción del neurotransmisor con el receptor

22
Sinapsis exitatoria e inhibitoria

23
NEUROTRANSMISORES

24
Naturaleza química variada de los Neurotransmisores

25
Neurotransmisores

26
ACCIÓN DE LAS DROGAS EN LA SINAPSIS

27
Etapas de la sinapsis que pueden verse afectadas por drogas

28
Efectos de las encefalinas y morfina en el control del dolor (anestésicos)
")
29
Estimulación de la sinapsis por drogas

30
CONTRACCIÓN MUSCULAR

31
Tejido muscular: efectores con acción mecánica o motora
Introducción Tejido muscular: efectores con acción mecánica o motora Formado por células excitables y contráctiles. Tipos: - Esquelético, unido a los huesos: responsable del movimiento coordinado y voluntario - Liso de las paredes de las vísceras (estómago, intestino, vasos sanguíneos…): involuntario - Cardiaco: estriado e involuntario Los principios básicos de excitación y contracción son aplicables a los tres.
: involuntario. - Cardiaco: estriado e involuntario. Los principios básicos de excitación y contracción son aplicables a los tres.")
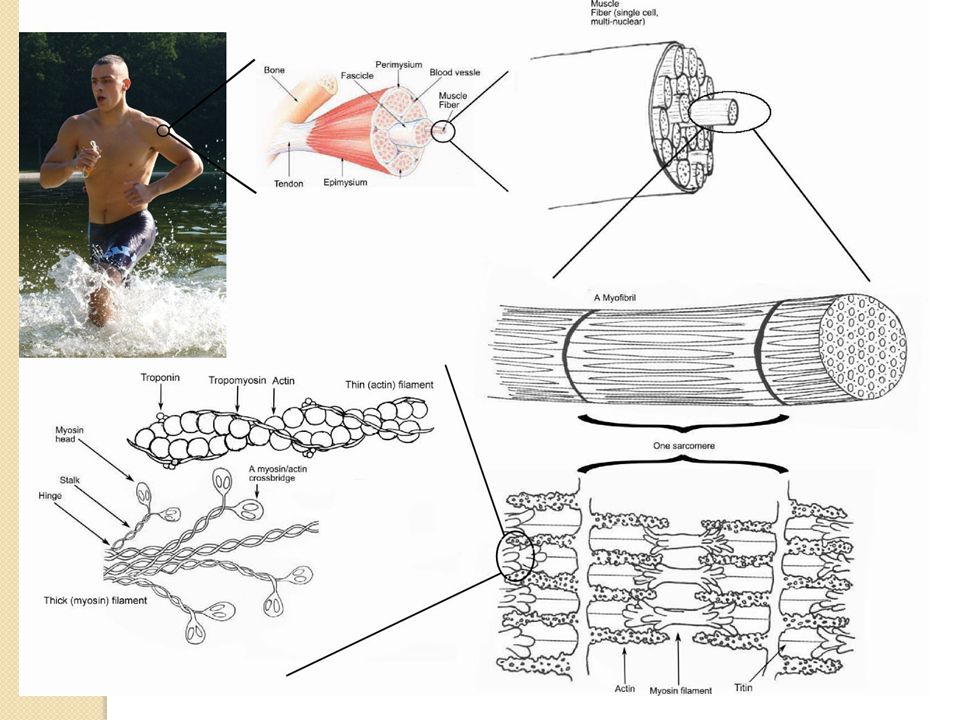
33
Músculo esquelético: características
Tejido muscular estriado rodeado de una vaina de tejido conectivo (epimisio) que lo inserta en los huesos (tendones). El músculo se divide en fascículos, y estos en fibras rodeadas de membrana plasmática con centenares o miles de miofibrillas que contienen los filamentos contráctiles (actina y miosina). Las estrías se deben a la disposición organizada de filamentos gruesos (miosina) y finos (actina). El sarcómero es la unidad contráctil del músculo esquelético. Estructura del músculo esquelético El tejido conjuntivo de los tendones rodea al músculo en una vaina llamada epimisio. Esta cubierta externa se extiende hacia el interior y divide al músculo en fascículos o columnas, de forma que cada uno de esos fascículos se encuentra rodeado por su propia vaina de conjuntivo denominadas perimisio. La disección de un fascículo muscular nos revela que se encuentra constituido por numerosas fibras musculares o miofibrillas. Cada miofibrilla está rodeada por una membrana celular denominada sarcolema que a su vez está recubierta por una fina capa de conjuntivo llamada endomisio. De esta forma es muy fácil recordar la estructura de músculo como si fuera la célula muscular un cigarrillo que sed encuentra en un paquete de cigarros que a su vez se guarda en un cartón. Las células musculares poseen los mismos orgánulos que el resto de células del organismo, sin embargo poseen una diferencia respecto la mayoría de células del cuerpo. Son multinucleadas. Esta estructura se debe a que cada fibra muscular está formada por la unión de varios mioblastos embrionarios. Otra característica notable de las fibras musculares esqueléticas es su aspecto estriado al microscopio.
 que lo inserta en los huesos (tendones). El músculo se divide en fascículos, y estos en fibras rodeadas de membrana plasmática con centenares o miles de miofibrillas que contienen los filamentos contráctiles (actina y miosina). Las estrías se deben a la disposición organizada de filamentos gruesos (miosina) y finos (actina). El sarcómero es la unidad contráctil del músculo esquelético. Estructura del músculo esquelético. El tejido conjuntivo de los tendones rodea al músculo en una vaina llamada epimisio. Esta cubierta externa se extiende hacia el interior y divide al músculo en fascículos o columnas, de forma que cada uno de esos fascículos se encuentra rodeado por su propia vaina de conjuntivo denominadas perimisio. La disección de un fascículo muscular nos revela que se encuentra constituido por numerosas fibras musculares o miofibrillas. Cada miofibrilla está rodeada por una membrana celular denominada sarcolema que a su vez está recubierta por una fina capa de conjuntivo llamada endomisio. De esta forma es muy fácil recordar la estructura de músculo como si fuera la célula muscular un cigarrillo que sed encuentra en un paquete de cigarros que a su vez se guarda en un cartón. Las células musculares poseen los mismos orgánulos que el resto de células del organismo, sin embargo poseen una diferencia respecto la mayoría de células del cuerpo. Son multinucleadas. Esta estructura se debe a que cada fibra muscular está formada por la unión de varios mioblastos embrionarios. Otra característica notable de las fibras musculares esqueléticas es su aspecto estriado al microscopio.")
34
Músculo esquelético: características
Banda A: filamentos de miosina solapados con los de actina Banda I: filamentos de actina que parten del disco Z Banda H: filamentos de miosina sin solapamiento con los de actina Características fibra (célula) muscular: - Membrana plasmática = sarcolema Multinucleada Retículo endoplásmico muy desarrollado (= sarcoplásmico) Gran cantidad de mitocondrias Podemos observar como al microscopio se ven unas bandas claras denominadas bandas I y unas bandas oscuras denominadas bandas A. Al microscopio electrónico en la parte media de las bandas I se observan unas líneas oscuras denominadas discos Z. El origen de las bandas radica en 2 tipos de filamentos unos gruesos, son los filamentos de miosina y dan lugar a la banda A y unos finos, filamentos de actina que dan lugar a la banda clara I. Detalladamente, desde el disco Z, el cual cruza toda la miofibrilla y también cruza de una miofibrilla a otra, parten filamentos de actina en ambas direcciones dando lugar a unas bandas claras. Estos filamentos de actina se intercalan y se solapan con los de miosina, dando lugar a las bandas oscuras A. Las regiones centrales más claras de las bandas A se denominan bandas H (de Helle, palabra alemana que significa brillante), así pues las banda H sólo contienen filamentos gruesos de miosina sin solapamiento con los filamentos finos. Cuando sólo hay miosina se recupera la apariencia clara hasta la región central de la miosina, lugar donde se establecen puentes entre las miosinas. Este aspecto estriado es típico tanto de músculo esquelético como de músculo cardíaco. La porción de la miofibrilla localizada entre dos discos Z se denomina sarcómero. Para no inducir a error se muestra, además de la visión lateral, una visión tridimensional de la miofibrilla. En cada miofibrilla existen numerosos sarcómeros fuera del plano de corte y de esta forma podemos observar los discos Z y que los filamentos finos que atraviesan estos discos Z rodean a los filamentos gruesos con una disposición hexagonal. Hacia el final del tema estudiaremos las características de la inervación muscular. Aquí únicamente me gustaría recalcar que cada fibra muscular estriada recibe una única terminación axónica procedente de una neurona motora somática. (DEFINIR CONCRETAMENTE UNIDAD MOTORA).
 muscular: - Membrana plasmática = sarcolema. Multinucleada. Retículo endoplásmico muy desarrollado (= sarcoplásmico) Gran cantidad de mitocondrias. Podemos observar como al microscopio se ven unas bandas claras denominadas bandas I y unas bandas oscuras denominadas bandas A. Al microscopio electrónico en la parte media de las bandas I se observan unas líneas oscuras denominadas discos Z. El origen de las bandas radica en 2 tipos de filamentos unos gruesos, son los filamentos de miosina y dan lugar a la banda A y unos finos, filamentos de actina que dan lugar a la banda clara I. Detalladamente, desde el disco Z, el cual cruza toda la miofibrilla y también cruza de una miofibrilla a otra, parten filamentos de actina en ambas direcciones dando lugar a unas bandas claras. Estos filamentos de actina se intercalan y se solapan con los de miosina, dando lugar a las bandas oscuras A. Las regiones centrales más claras de las bandas A se denominan bandas H (de Helle, palabra alemana que significa brillante), así pues las banda H sólo contienen filamentos gruesos de miosina sin solapamiento con los filamentos finos. Cuando sólo hay miosina se recupera la apariencia clara hasta la región central de la miosina, lugar donde se establecen puentes entre las miosinas. Este aspecto estriado es típico tanto de músculo esquelético como de músculo cardíaco. La porción de la miofibrilla localizada entre dos discos Z se denomina sarcómero. Para no inducir a error se muestra, además de la visión lateral, una visión tridimensional de la miofibrilla. En cada miofibrilla existen numerosos sarcómeros fuera del plano de corte y de esta forma podemos observar los discos Z y que los filamentos finos que atraviesan estos discos Z rodean a los filamentos gruesos con una disposición hexagonal. Hacia el final del tema estudiaremos las características de la inervación muscular. Aquí únicamente me gustaría recalcar que cada fibra muscular estriada recibe una única terminación axónica procedente de una neurona motora somática. (DEFINIR CONCRETAMENTE UNIDAD MOTORA).")
35
Filamentos contráctiles: miosina
Filamentos gruesos compuestos por múltiples moléculas de miosina (200 o más). Formada por 2 cadenas pesadas formando una doble hélice (cola de la molécula de miosina) y 4 cadenas ligeras (cabeza de miosina). La cabeza está separada de la hélice mediante un brazo flexible. El conjunto cabeza-brazo se llama puente cruzado y participa directamente en la contracción. La cabeza de miosina posee actividad ATPasa y puede unirse a la actina. Los filamentos de miosina están compuestos por múltiples moléculas de miosina. Estas moléculas interaccionan con por uno de sus lados con los filamentos de actina. La molécula de miosina está formada por seis cadenas polipeptídicas, 2 cadenas pesadas de y cuatro cadenas ligeras de Las dos pesadas se enrollan en espiral para formar una doble hélice, denominada cola de la molécula de miosina. Un extremo de cada una de esas cadenas esta plegado en una estructura polipeptídica globulosa denominado cabeza de miosina. Así existen dos cabezas libres situadas una al lado de la otra en un extremo de la cadena de miosina. En estas cabezas se localizan las cadenas ligeras, 2 por cada cabeza y ayudan a controlar la función de la cabeza durante la contracción muscular. Un filamento de miosina está constituido por 200 o mas moléculas individuales de miosina, en la diapositiva se observa como se agrupan para formar el cuerpo del filamento. Cada cabeza se encuentra separada de la porción de hélice mediante un brazo. Al conjunto cabeza-brazo se le denomina puente cruzado. El puente cruzado es flexible y participa directamente en el proceso real de contracción. Se puede observar como en el centro de la fibra no existen cabezas de puente cruzado ya que se extienden alejándose de el centro. Además la fibra de miosina se retuerce sobre sí misma, de modo que cada par sucesivo de puentes cruzados está desplazado axialmente del conjunto previo en 120grados. Así se asegura que los puentes cruzados se extiendan en todas las direcciones alrededor del filamento. Otra característica de la cabeza de miosina es su actividad ATPasa, esencial como veremos para la contracción muscular.
. Formada por 2 cadenas pesadas formando una doble hélice (cola de la molécula de miosina) y 4 cadenas ligeras (cabeza de miosina). La cabeza está separada de la hélice mediante un brazo flexible. El conjunto cabeza-brazo se llama puente cruzado y participa directamente en la contracción. La cabeza de miosina posee actividad ATPasa y puede unirse a la actina. Los filamentos de miosina están compuestos por múltiples moléculas de miosina. Estas moléculas interaccionan con por uno de sus lados con los filamentos de actina. La molécula de miosina está formada por seis cadenas polipeptídicas, 2 cadenas pesadas de y cuatro cadenas ligeras de Las dos pesadas se enrollan en espiral para formar una doble hélice, denominada cola de la molécula de miosina. Un extremo de cada una de esas cadenas esta plegado en una estructura polipeptídica globulosa denominado cabeza de miosina. Así existen dos cabezas libres situadas una al lado de la otra en un extremo de la cadena de miosina. En estas cabezas se localizan las cadenas ligeras, 2 por cada cabeza y ayudan a controlar la función de la cabeza durante la contracción muscular. Un filamento de miosina está constituido por 200 o mas moléculas individuales de miosina, en la diapositiva se observa como se agrupan para formar el cuerpo del filamento. Cada cabeza se encuentra separada de la porción de hélice mediante un brazo. Al conjunto cabeza-brazo se le denomina puente cruzado. El puente cruzado es flexible y participa directamente en el proceso real de contracción. Se puede observar como en el centro de la fibra no existen cabezas de puente cruzado ya que se extienden alejándose de el centro. Además la fibra de miosina se retuerce sobre sí misma, de modo que cada par sucesivo de puentes cruzados está desplazado axialmente del conjunto previo en 120grados. Así se asegura que los puentes cruzados se extiendan en todas las direcciones alrededor del filamento. Otra característica de la cabeza de miosina es su actividad ATPasa, esencial como veremos para la contracción muscular.")
36
Filamentos contráctiles: actina
Filamentos finos constituidos por: doble hebra de actina, tropomiosina y troponina. La tropomiosina se enrolla en espiral alrededor de la actina. En reposo impide atracción entre los filamentos de actina y de miosina. La troponina (complejo de) se une a los lados de la tropomiosina. La troponina I posee gran afinidad por la actina, la T por la tropomiosina y la C por el calcio. Filamento de actina está constituido por actina, tropomiosina y troponina. Una doble hebra de filamento de actina se enrolla sobre sí misma en una hélice. Cada filamento de esta actina F de la doble hélice está compuesta por actina G polimerizada. Unida a cada una de las moléculas de actina G se encuentra una molécula de ADP. Se cree que estas moléculas de ADP son los sitios activos de los filamentos de actina con los que interactúan los puentes cruzados de los filamentos de miosina para producir la contracción muscular. Las moléculas de tropomiosina se enrollan en espiral alrededor de los lados de la hélice de actina F. En estado de reposo las moléculas de tropomiosina descansan sobre los sitios activos de las hebras de actina, por lo que no puede haber atracción entre los filamentos de actina y de miosina para producir la contracción. La troponina se encuentra unida a los lados de las moléculas de tropomiosina. En realidad son complejos de tres subunidades proteicas unidas de forma laxa, cada una de las cuales tiene un papel específico en el control de la contracción muscular. La troponina I posee gran afinidad por la actina, la T por la tropomiosina y la C por los iones calcio. Se cree que este complejo une la tropomiosina a la actina, y que la fuerte afinidad de la troponina por los iones calcio inicia el proceso de contracción. La relación lado a lado que existe entre los filamentos de actina y miosina no es fácil de mantener. Esto se logra mediante la acción de un gran número de moléculas filamentosas de una proteína llamada titina. La proteína titina es una de las proteínas más grandes del organismo (tiene un PM de ). Debido a su naturaleza filamentosa es una proteína muy elástica por lo que actúa como armazón que reviste los filamentos de actina y miosina para construir la maquinaria contráctil de trabajo del sarcómero.
 se une a los lados de la tropomiosina. La troponina I posee gran afinidad por la actina, la T por la tropomiosina y la C por el calcio. Filamento de actina está constituido por actina, tropomiosina y troponina. Una doble hebra de filamento de actina se enrolla sobre sí misma en una hélice. Cada filamento de esta actina F de la doble hélice está compuesta por actina G polimerizada. Unida a cada una de las moléculas de actina G se encuentra una molécula de ADP. Se cree que estas moléculas de ADP son los sitios activos de los filamentos de actina con los que interactúan los puentes cruzados de los filamentos de miosina para producir la contracción muscular. Las moléculas de tropomiosina se enrollan en espiral alrededor de los lados de la hélice de actina F. En estado de reposo las moléculas de tropomiosina descansan sobre los sitios activos de las hebras de actina, por lo que no puede haber atracción entre los filamentos de actina y de miosina para producir la contracción. La troponina se encuentra unida a los lados de las moléculas de tropomiosina. En realidad son complejos de tres subunidades proteicas unidas de forma laxa, cada una de las cuales tiene un papel específico en el control de la contracción muscular. La troponina I posee gran afinidad por la actina, la T por la tropomiosina y la C por los iones calcio. Se cree que este complejo une la tropomiosina a la actina, y que la fuerte afinidad de la troponina por los iones calcio inicia el proceso de contracción. La relación lado a lado que existe entre los filamentos de actina y miosina no es fácil de mantener. Esto se logra mediante la acción de un gran número de moléculas filamentosas de una proteína llamada titina. La proteína titina es una de las proteínas más grandes del organismo (tiene un PM de ). Debido a su naturaleza filamentosa es una proteína muy elástica por lo que actúa como armazón que reviste los filamentos de actina y miosina para construir la maquinaria contráctil de trabajo del sarcómero.")
37
Contracción muscular = Disminución en la longitud de las fibras individuales. Disminución en la distancia entre los discos Z sin acortamiento de las bandas A. Las bandas I disminuyen de longitud. La disminución de longitud del sarcómero se debe al deslizamiento de los filamentos finos sobre y entre los filamentos gruesos. La contracción muscular produce una disminución en la longitud de sus fibras individuales. Este acortamiento se debe a la disminución de tamaño en las miofobrillas que por su parte está causada por la disminución en la distancia entre cada dos discos Z. En esta aproximación entre los discos Z, las bandas A no presentan un acortamiento, sino que se muestra una aproximación entre las bandas A y que las bandas I, que representan la distancia entre las bandas A de los sarcómeros, disminuyen de longitud. De todos modos y aunque se muestre una disminución de su longitud no se produce un acortamiento de la banda I. Podemos observar como el acortamiento de los sarcómeros se debe al deslizamiento de los filamentos finos sobre y entre los filamentos gruesos.

38
Contracción muscular = Disminución en la longitud de los sarcómeros y por tanto de las fibras musculares. Las bandas A no varían, mientras que las bandas I se estrechan. La contracción muscular produce una disminución en la longitud de sus fibras individuales. Este acortamiento se debe a la disminución de tamaño en las miofobrillas que por su parte está causada por la disminución en la distancia entre cada dos discos Z. En esta aproximación entre los discos Z, las bandas A no presentan un acortamiento, sino que se muestra una aproximación entre las bandas A y que las bandas I, que representan la distancia entre las bandas A de los sarcómeros, disminuyen de longitud. De todos modos y aunque se muestre una disminución de su longitud no se produce un acortamiento de la banda I. Podemos observar como el acortamiento de los sarcómeros se debe al deslizamiento de los filamentos finos sobre y entre los filamentos gruesos.

39
Contracción muscular Troponina y tropomiosina regulan la unión de los puentes actina-miosina. En reposo, la tropomiosina bloquea la unión de los puentes cruzados a la actina. El desplazamiento de la tropomiosina requiere la interacción de la troponina con Ca2+ liberado por el RS. Este desplazamiento muestra los puntos activos de la actina. La ATPasa de la miosina hidroliza el ATP a ADP y Pi, que se mantienen unidos a la cabeza. Puentes cruzados de miosina se unen a las moléculas de actina. Las cabezas de miosina se inclinan al liberar el Pi, provocando el deslizamiento sobre la actina (golpe de fuerza). El ADP ha de ser sustituido por un nuevo ATP para que la cabeza se separe de la actina Una vez finalizado el estímulo nervioso bombas de Ca2+ devuelven el catión al RS. Al separar El Ca2+ la tropomiosina vuelve a su sitio cubriendo los puntos activos de la actina: relajación Este mecanismo de deslizamiento que se da en la contracción muscular se debe a la acción de los puentes cruzados que se proyectan desde la miosina hacia la actina. Como ya hemos indicado estos puentes cruzados se extienden desde el eje de los filamentos gruesos formando unos brazos que finalizan en las cabezas globulares. Recordar que cada miosina presenta dos cabezas globulares que sirven como puentes cruzados y que la orientación de las cabezas de miosina a cada lado de un sarcómero es opuesta a la de las cabezas del otro lado de manera que cuando las cabezas de miosina establecen puentes cruzados con la actina de cada lado del sarcómero tiran de la actina de cada lado hacia el centro. Se ha observado como la presencia de un filamento de actina pura, sin la presencia del complejo troponina-tropomiosina, se une de forma instantánea y fuertemente con las cabezas de las moléculas de miosina en presencia de iones magnesio y de ATP (ambos abundantes en la miofibrilla). Si a este filamento se le añade el complejo troponina-tropomiosina, esta unión no tiene lugar. Por lo que se cree que los sitios activos de unión de la actina normal en músculo relajado están inhibidos o recubiertos físicamente por el complejo troponina-tropomiosina. Por ello, en el músculo relajado, los sitios no se pueden unir a las cabezas de los filamentos de miosina para producir la contracción. Cada cabeza globular de miosina de un puente cruzado contiene un lugar de unión a ATP asociado de manera estrecha al lugar de unión a la actina. La actividad ATPasa de las cabezas de miosina activa a las misma mediante la degradación del ATP. El ADP y Pi generado quedan unidos a las cabezas de miosina hasta que se han unido a los sitios activos de unión de la actina. Cuando se establece el puente cruzado se libera el Pi y se produce una modificación de la configuración de la proteína de miosina causando un golpe de fuerza que tira de los filamentos finos hacia el centro de las bandas A. El ADP se libera cuando los puentes cruzados unen otra molécula de ATP al final del golpe de fuerza. La liberación de ADP por la unión de ATP es necesaria para que se rompa la unión establecida entre la actina y la miosina al final del golpe de fuerza. LA hidrólisis de esta molécula de ATP por la actividad ATPasa de la cabeza de miosina la activará de nuevo. Es muy importante el hecho que antes de que el puente cruzado se pueda unir a la actina la molécula de ATP debe ser hidrolizada y que para que el puente cruzado se libere de la actina debe unirse otra nueva molécula de ATP. Por ejemplo, en el caso de la rigidez cadavérica, tiene lugar debido a la ausencia de ATP cuando muere el músculo. Sin ATP, el ADP queda unido a los puentes cruzados y estos permanecen estrechamente unidos a la actina. Los puentes cruzados son muy cortos, por ello un único ciclo de contracción con golpe de fuerza de todos los puentes cruzados del músculo, sólo lo acorta en un 1% respecto a la situación de reposo. Sin embargo se ha observado como los músculos pueden llegar a presentar un acortamiento del 60%, por lo tanto es obvio que se debe repetir muchas veces los ciclos de contracción. Así que los puentes cruzados se separan de la aactina al final del golpe de fuerza, vuelven a presentar su conveniente orientación de reposo y se vuelven a unir a la actina para repetir el ciclo. Además, hay que pensar que durante la contracción muscular normal sólo están unidos una parte de los puentes cruzados en cada momento, es decir, no presentan un sincronismo como ocurre con los componentes de un equipo de remo, sino que se produce un estiramiento asincrónico como ocurre con la fuerza ejercida en una competición de estiramiento de cuerda o la fuerza ejercida por cuatro personas al empujar un coche se genera una fuerza constante pero no todos la aplican simultáneamente. Para que el músculo se relaje se debe impedir la unión de los puentes cruzados de la miosina a la actina. Se ha indicado anteriormente el papel que desempeña el complejo Troponina-tropomiosina como inhibidores de la unión a los sitios activos de la actina. La actina (actina F) es un polímero formado por unas subunidades globulares (actina G) dispuestas en doble hélice. En el surco que queda entre la doble fila de monómeros de actina se dispone la tropomiosina. La troponina, con sus tres subunidades, actúa conjuntamente con la tropomiosina regulando la unión de los puentes cruzados. En el músculo relajado, la tropomiosina ocupa una posición en los filamentos finos que le permite bloquear físicamente la unión de los puentes cruzados a sus lugares específicos de fijación en la actina. Así para que los puentes cruzados de la miosina puedan unirse a la actina, se debe desplazar la tropomiosina. Este desplazamiento requiere la interacción de la troponina con Ca2+. En un músculo relajado, cuando la tropomiosina se encuentra bloqueando los sitios de unión de los puentes cruzados a la actina la concentración de Ca2+ en el sarcoplasma (citoplasma de la célula muscular) es muy baja. Al realizarse el estímulo sobre la célula muscular para que se contraiga se ponen en marcha diversos mecanismos que aumentan la concentración de ca2+ en el sarcoplasma. Parte de ese Ca2+ se une a la troponina lo cual origina un cambio conformacional que hace que la tropomiosina unida<a la troponina se desplacen y se expongan los sitios de unión de la actina a los puentes cruzados. Inmediatamente se produce esta unión, los golpes de fuerza y la contracción muscular. Así pues la posición de los complejos troponina-tropomiosina es ajustable, es decir, cuando el Ca2+ no está unido a la troponina, la tropomiosina permanece en una posición que inhibe la unión de los puentes cruzados a la actina y, por tanto, inhibe la contracción muscular. La unión de Ca2+ la troponina induce un cambio conformacional del complejo que expone los sitios activos de unión. Los ciclos de contracción que se generan pueden continuar siempre y cuando el Ca2+ permanezca unido a la troponina.
. El ADP ha de ser sustituido por un nuevo ATP para que la cabeza se separe de la actina. Una vez finalizado el estímulo nervioso bombas de Ca2+ devuelven el catión al RS. Al separar El Ca2+ la tropomiosina vuelve a su sitio cubriendo los puntos activos de la actina: relajación. Este mecanismo de deslizamiento que se da en la contracción muscular se debe a la acción de los puentes cruzados que se proyectan desde la miosina hacia la actina. Como ya hemos indicado estos puentes cruzados se extienden desde el eje de los filamentos gruesos formando unos brazos que finalizan en las cabezas globulares. Recordar que cada miosina presenta dos cabezas globulares que sirven como puentes cruzados y que la orientación de las cabezas de miosina a cada lado de un sarcómero es opuesta a la de las cabezas del otro lado de manera que cuando las cabezas de miosina establecen puentes cruzados con la actina de cada lado del sarcómero tiran de la actina de cada lado hacia el centro. Se ha observado como la presencia de un filamento de actina pura, sin la presencia del complejo troponina-tropomiosina, se une de forma instantánea y fuertemente con las cabezas de las moléculas de miosina en presencia de iones magnesio y de ATP (ambos abundantes en la miofibrilla). Si a este filamento se le añade el complejo troponina-tropomiosina, esta unión no tiene lugar. Por lo que se cree que los sitios activos de unión de la actina normal en músculo relajado están inhibidos o recubiertos físicamente por el complejo troponina-tropomiosina. Por ello, en el músculo relajado, los sitios no se pueden unir a las cabezas de los filamentos de miosina para producir la contracción. Cada cabeza globular de miosina de un puente cruzado contiene un lugar de unión a ATP asociado de manera estrecha al lugar de unión a la actina. La actividad ATPasa de las cabezas de miosina activa a las misma mediante la degradación del ATP. El ADP y Pi generado quedan unidos a las cabezas de miosina hasta que se han unido a los sitios activos de unión de la actina. Cuando se establece el puente cruzado se libera el Pi y se produce una modificación de la configuración de la proteína de miosina causando un golpe de fuerza que tira de los filamentos finos hacia el centro de las bandas A. El ADP se libera cuando los puentes cruzados unen otra molécula de ATP al final del golpe de fuerza. La liberación de ADP por la unión de ATP es necesaria para que se rompa la unión establecida entre la actina y la miosina al final del golpe de fuerza. LA hidrólisis de esta molécula de ATP por la actividad ATPasa de la cabeza de miosina la activará de nuevo. Es muy importante el hecho que antes de que el puente cruzado se pueda unir a la actina la molécula de ATP debe ser hidrolizada y que para que el puente cruzado se libere de la actina debe unirse otra nueva molécula de ATP. Por ejemplo, en el caso de la rigidez cadavérica, tiene lugar debido a la ausencia de ATP cuando muere el músculo. Sin ATP, el ADP queda unido a los puentes cruzados y estos permanecen estrechamente unidos a la actina. Los puentes cruzados son muy cortos, por ello un único ciclo de contracción con golpe de fuerza de todos los puentes cruzados del músculo, sólo lo acorta en un 1% respecto a la situación de reposo. Sin embargo se ha observado como los músculos pueden llegar a presentar un acortamiento del 60%, por lo tanto es obvio que se debe repetir muchas veces los ciclos de contracción. Así que los puentes cruzados se separan de la aactina al final del golpe de fuerza, vuelven a presentar su conveniente orientación de reposo y se vuelven a unir a la actina para repetir el ciclo. Además, hay que pensar que durante la contracción muscular normal sólo están unidos una parte de los puentes cruzados en cada momento, es decir, no presentan un sincronismo como ocurre con los componentes de un equipo de remo, sino que se produce un estiramiento asincrónico como ocurre con la fuerza ejercida en una competición de estiramiento de cuerda o la fuerza ejercida por cuatro personas al empujar un coche se genera una fuerza constante pero no todos la aplican simultáneamente. Para que el músculo se relaje se debe impedir la unión de los puentes cruzados de la miosina a la actina. Se ha indicado anteriormente el papel que desempeña el complejo Troponina-tropomiosina como inhibidores de la unión a los sitios activos de la actina. La actina (actina F) es un polímero formado por unas subunidades globulares (actina G) dispuestas en doble hélice. En el surco que queda entre la doble fila de monómeros de actina se dispone la tropomiosina. La troponina, con sus tres subunidades, actúa conjuntamente con la tropomiosina regulando la unión de los puentes cruzados. En el músculo relajado, la tropomiosina ocupa una posición en los filamentos finos que le permite bloquear físicamente la unión de los puentes cruzados a sus lugares específicos de fijación en la actina. Así para que los puentes cruzados de la miosina puedan unirse a la actina, se debe desplazar la tropomiosina. Este desplazamiento requiere la interacción de la troponina con Ca2+. En un músculo relajado, cuando la tropomiosina se encuentra bloqueando los sitios de unión de los puentes cruzados a la actina la concentración de Ca2+ en el sarcoplasma (citoplasma de la célula muscular) es muy baja. Al realizarse el estímulo sobre la célula muscular para que se contraiga se ponen en marcha diversos mecanismos que aumentan la concentración de ca2+ en el sarcoplasma. Parte de ese Ca2+ se une a la troponina lo cual origina un cambio conformacional que hace que la tropomiosina unida<a la troponina se desplacen y se expongan los sitios de unión de la actina a los puentes cruzados. Inmediatamente se produce esta unión, los golpes de fuerza y la contracción muscular. Así pues la posición de los complejos troponina-tropomiosina es ajustable, es decir, cuando el Ca2+ no está unido a la troponina, la tropomiosina permanece en una posición que inhibe la unión de los puentes cruzados a la actina y, por tanto, inhibe la contracción muscular. La unión de Ca2+ la troponina induce un cambio conformacional del complejo que expone los sitios activos de unión. Los ciclos de contracción que se generan pueden continuar siempre y cuando el Ca2+ permanezca unido a la troponina.")
40
Contracción muscular Rigor mortis: sin el ATP producido por el metabolismo celular, el ADP queda unido a la cabeza de miosina, y ésta queda “enganchada” a la actina.

41
Contracción muscular: retículo sarcoplásmico
Sarcolema = membrana plasmática Retículo sarcoplásmico = retículo endoplasmático especializado a modo de cisternas donde se almacena Ca2+ : su concentración es muy baja en el citoplasma. Túbulos T = invaginaciones del sarcolema hacia el interior celular que hacen llegar el potencial de acción a toda la fibra muscular Como se ha indicado la contracción muscular se produce siempre y cuando se una a la troponina una cantidad suficiente de Ca2+, lo cual ocurre cuando en el sarcoplasma se alcanzan valores superiores a 10e-6M. Y por supuesto que para que se produzca la relajación muscular los niveles de Ca2+ en el sarcoplasma deben ser menores a dicha concentración. En la relajación muscular se transporta activamente el Ca2+ fuera del sarcoplasma hacia el retículo sarcoplásmico. El retículo sarcoplásmico es un tipo modificado de retículo endoplasmático constituido por cisternas y por conductos interconectados que rodean a cada miofibrilla en el interior de la célula muscular. Las cisternas terminales, la primera porción mencionada, están en contacto con los túbulos transversales o túbulos T, que corren a lo largo de las líneas Z que limitan el sarcómero. (Libro histología). Por tanto los túbulos T se encuentran separando las cisternas terminales. Entos estrechos túneles membranosos se forman a partir del sarcolema (membrana celular muscular) y muestran continuidad con el mismo. Así pues, los túbulos T se abren hacia el espacio extracelular a través de poros localizados en la superficie de la célula y tienen capacidad de conducir potenciales de acción. En la célula muscular relajada la mayor parte del Ca2+ permanece almacenado en las cisternas terminales. Cuando se produce la estimulación de la fibra muscular para que se contraiga ya sea en respuesta a una neurona motora in vivo o al efecto de un choque eléctrico in vitro, el Ca2+ almacenado se libera del retículo sarcoplásmico y puede unirse a la troponina. Al interrumpirse la estimulación de la fibra muscular el Ca2+ es devuelto al retículo sarcoplásmico mediante transporte activo.
. Por tanto los túbulos T se encuentran separando las cisternas terminales. Entos estrechos túneles membranosos se forman a partir del sarcolema (membrana celular muscular) y muestran continuidad con el mismo. Así pues, los túbulos T se abren hacia el espacio extracelular a través de poros localizados en la superficie de la célula y tienen capacidad de conducir potenciales de acción. En la célula muscular relajada la mayor parte del Ca2+ permanece almacenado en las cisternas terminales. Cuando se produce la estimulación de la fibra muscular para que se contraiga ya sea en respuesta a una neurona motora in vivo o al efecto de un choque eléctrico in vitro, el Ca2+ almacenado se libera del retículo sarcoplásmico y puede unirse a la troponina. Al interrumpirse la estimulación de la fibra muscular el Ca2+ es devuelto al retículo sarcoplásmico mediante transporte activo.")
42
Unión neuromuscular El músculo esquelético está inervado por grandes fibras mielinizadas originadas en las motoneuronas de la médula espinal. Las fibras nerviosas se ramifican e inervan entre 3 y varios cientos de fibras musculares. En los movimientos finos una motoneurona inerva pocas fibras musculares. Unidad motora: conjunto de fibras musculares inervadas por una sola motoneurona. La unión neuromuscular, cerca del punto medio de la fibra muscular, se llama placa motora terminal. Las fibras de músculo esquelético están inervadas por fibras nerviosas grandes y mielinizadas originadas en las grandes motoneuronas del asta anterior de la médula espinal. En el músculo cada fibra nerviosa se ramifica y puede estimular entre 3 y varios cientos de fibras musculares esqueléticas (así por ejemplo en el ojo hay una fibra nerviosa por cada fibra muscular, se precisa mucha precisión de movimiento; si un músculo no ha de realizar movimientos delicados la fibra nerviosa se encuentra muy ramificada). Cada terminación nerviosa establece una unión denominada unión neuromuscular localizada cerca del punto medio de la fibra muscular, en un punto que se ha denominado placa motora terminal, de este modo cuando le llega un potencial de acción se transmite en ambas direcciones hacia los extremos. Normalmente sólo existe una de estas uniones por fibra muscular (excepciones aprox un 2%). Antes de descubrirse el microscopio electrónico se aceptó que los axones al acercarse a la fibra muscular y antes de ramificarse en las ramas terminales, pierde su vaina de mielina. Las ramitas se acercan a la superficie de la fibra muscular únicamente cubiertas por la vaina de la células de schwann. Gracias al desarrollo tecnológico de la microscopía electrónica se pudieron observar muchos más detalles. Se observó una invaginación del sarcolema a la que se denominó canal o depresión sináptica y un espacio que separaba la fibra nerviosa de la muscular, es decir un espacio o hendidura sináptica. En el fondo del canal existen numerosos pliegues del sarcolema que aumentan la superficie de la célula muscular expuesta al neurotransmisor liberado por la fibra nerviosa, los cuales se denominan hendiduras subneurales. Además en el terminal nervioso se observaron muchas mitocondrias, indicativo de una elevada tasa metabólica, ya que suministran ATP, imprescindible para la síntesis de acetilcolina. La acetilcolina como se indicó en el tema anterior se sintetiza en el citoplasma del terminal y se absorbe rápidamente en las vesículas sinápticas. En la hendidura sináptica existen acetilcolinesterasas que hidrolizan la acetilcolina rápidamente tras ser liberadas de las vesículas. Como vimos en el tema anterior los receptores nicotinicos de acetilcolina son en realidad canales iónicos regulados por acetilcolina. De este modo cuando el potencial de acción de la fibra nerviosa llega al botón axónico se estimula la liberación de acetilcolina, la cual, mediante exocitosis es vertida a la hendidura sináptica. Parte de esa acetilcolina llegará a sus receptores en la célula muscular, localizados en las bocas de las hendiduras subneurales localizadas próximas a los llugares de liberación de acetilcolina. Como ya se indicó la unión de dos moléculas de acetilcolina abre el canal y se permite el paso de Na+, K+ y Ca2+, aunque el que entra mayoritariamente a favor de gradiente de concentración es el Na+. Los iones negativos no pasan por la existencia de cargas negativas en la boca del canal. La entrada de iones positivos origina un potencial postsináptico (potencial local) denominado potencial de la placa terminal, y se genera una despolarización que iniciará un potencial de acción que originará la contracción muscular.
. Cada terminación nerviosa establece una unión denominada unión neuromuscular localizada cerca del punto medio de la fibra muscular, en un punto que se ha denominado placa motora terminal, de este modo cuando le llega un potencial de acción se transmite en ambas direcciones hacia los extremos. Normalmente sólo existe una de estas uniones por fibra muscular (excepciones aprox un 2%). Antes de descubrirse el microscopio electrónico se aceptó que los axones al acercarse a la fibra muscular y antes de ramificarse en las ramas terminales, pierde su vaina de mielina. Las ramitas se acercan a la superficie de la fibra muscular únicamente cubiertas por la vaina de la células de schwann. Gracias al desarrollo tecnológico de la microscopía electrónica se pudieron observar muchos más detalles. Se observó una invaginación del sarcolema a la que se denominó canal o depresión sináptica y un espacio que separaba la fibra nerviosa de la muscular, es decir un espacio o hendidura sináptica. En el fondo del canal existen numerosos pliegues del sarcolema que aumentan la superficie de la célula muscular expuesta al neurotransmisor liberado por la fibra nerviosa, los cuales se denominan hendiduras subneurales. Además en el terminal nervioso se observaron muchas mitocondrias, indicativo de una elevada tasa metabólica, ya que suministran ATP, imprescindible para la síntesis de acetilcolina. La acetilcolina como se indicó en el tema anterior se sintetiza en el citoplasma del terminal y se absorbe rápidamente en las vesículas sinápticas. En la hendidura sináptica existen acetilcolinesterasas que hidrolizan la acetilcolina rápidamente tras ser liberadas de las vesículas. Como vimos en el tema anterior los receptores nicotinicos de acetilcolina son en realidad canales iónicos regulados por acetilcolina. De este modo cuando el potencial de acción de la fibra nerviosa llega al botón axónico se estimula la liberación de acetilcolina, la cual, mediante exocitosis es vertida a la hendidura sináptica. Parte de esa acetilcolina llegará a sus receptores en la célula muscular, localizados en las bocas de las hendiduras subneurales localizadas próximas a los llugares de liberación de acetilcolina. Como ya se indicó la unión de dos moléculas de acetilcolina abre el canal y se permite el paso de Na+, K+ y Ca2+, aunque el que entra mayoritariamente a favor de gradiente de concentración es el Na+. Los iones negativos no pasan por la existencia de cargas negativas en la boca del canal. La entrada de iones positivos origina un potencial postsináptico (potencial local) denominado potencial de la placa terminal, y se genera una despolarización que iniciará un potencial de acción que originará la contracción muscular.")
43
Acoplamiento excitación-contracción
El potencial de acción generado en la placa motora terminal se propaga por toda la fibra, y es conducido hacia el interios por los túbulos T, que están en contacto con el RS. La despolarización abre canales de Ca+2 voltaje dependientes. El aumento del Ca+2 en el citoplasma activa la contracción muscular. En las células musculares los potenciales de acción pueden conducirse hacia el interior de las fibras a través de la membrana de los túbulos transversales. En los túbulos T los potenciales de acción dan lugar a la liberación de Ca2+ a partir del retículo sarcoplásmico. Como no existe una continuidad física entre el retículo sarcoplásmico y los túbulos T se cree que entre ambos debe existir un acoplamiento directo entre los canales Ca2+ voltaje dependientes a través de los cuales se libera el Ca2+.

Presentaciones similares