Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Transcripción Para que la información genética almacenada en el DNA sea utilizada por los organismos se requiere que sean sintetizadas cadenas de RNA complementarias al DNA por medio de un proceso conocido como transcripción (de “scribere”=escribir y “trans”= pasar de uno a otro, en este caso de una escritura a otra).
.")
2
Transcripción Fue adoptado por la evolución para tener un mensajero capaz de llevar la información almacenada en el DNA de forma parcializada, ordenada y regulada. El resultado de la transcripción es una molécula de RNA llamada transcrito.

3
Transcritos Cuando sirve como patrón para la síntesis de proteínas se conoce como RNA mensajero o RNAm; Cuando constituye parte de la estructura de los ribosomas, como RNA ribosomal o RNAr; Cuando participa en la síntesis de proteínas como adaptador de los aminoácidos, como RNA de transferencia o RNAt; o Puede tener varias otras actividades como enzimáticas, ribozima o actividades de regulación de la expresión genética, RNA de interferencia o RNAi.

4
Transcripción y Superenrollamiento
RNA polimerasa Burbuja de Transcripción Enrollamiento Desenrollamiento 5' 3' 5' 3' 3' 5' Híbrido RNA-DNA RNA Dirección de la trancripción Superhélices positivas (b) Para transcribir una de las dos hebras de DNA, este se desenrolla transitoriamente. En todo momento están desenrollados unos 17 pares de bases. En la región desenrollada se encuentra un híbrido RNA-DNA corto (12 pares de bases). La burbuja de transcripción se desplaza tal como se muestra de izquierda a derecha manteniendo el ritmo con la síntesis de RNA. El DNA se desenrolla por delante y se vuelve a enrollar por detrás a medida que se transcribe el RNA. Las flechas indican la dirección en que rotan el DNA y el híbrido RNA-DNA para permitir este proceso. A medida que el DNA se vuelve a enrollar, se desplaza el híbrido RNA-DNA y se expulsa la hebra de RNA. (b) Superenrollamiento del DNA provocado por la transcripción. Se forman superhélices positivas por delante de la burbuja de transcripción y superhélices negativas por detrás de la misma. Superhélices negativas Dirección de la trancripción 5'
 Para transcribir una de las dos hebras de DNA, este se desenrolla transitoriamente. En todo momento están desenrollados unos 17 pares de bases. En la región desenrollada se encuentra un híbrido RNA-DNA corto (12 pares de bases). La burbuja de transcripción se desplaza tal como se muestra de izquierda a derecha manteniendo el ritmo con la síntesis de RNA. El DNA se desenrolla por delante y se vuelve a enrollar por detrás a medida que se transcribe el RNA. Las flechas indican la dirección en que rotan el DNA y el híbrido RNA-DNA para permitir este proceso. A medida que el DNA se vuelve a enrollar, se desplaza el híbrido RNA-DNA y se expulsa la hebra de RNA. (b) Superenrollamiento del DNA provocado por la transcripción. Se forman superhélices positivas por delante de la burbuja de transcripción y superhélices negativas por detrás de la misma. Superhélices. negativas. Dirección de la trancripción. 5")
5
Localización de transcritos
Transcritos de RNA DNA 36 x 103 pb La información de los adenovirus esta condificada en una molécula de DNA de doble hebra (36,000 pb), en la que ambas hebras contienen codificación de proteínas. La información para la mayor parte de proteína esta codificada en la hebra superior, pero parte está codificada en la hebra inferior y se transcribe en la dirección opuesta. La síntesis de los mRNA en este organismos es, de hecho más compleja. Muchos de los mensajeros se sintetizan incialmente en forma de un transcrito largo que proviene de dos tercios de la longitud del DNA. Este trasncrito es modificado posteriormente. Genoma de adenovirus, ambas hebras contienen codificación de proteínas. Muchos de los mensajeros se sintetizan inicialmente en forma de un transcrito largo que proviene de dos tercios de la longitud del DNA. Este transcrito es modificado posteriormente.
, en la que ambas hebras contienen codificación de proteínas. La información para la mayor parte de proteína esta codificada en la hebra superior, pero parte está codificada en la hebra inferior y se transcribe en la dirección opuesta. La síntesis de los mRNA en este organismos es, de hecho más compleja. Muchos de los mensajeros se sintetizan incialmente en forma de un transcrito largo que proviene de dos tercios de la longitud del DNA. Este trasncrito es modificado posteriormente. Genoma de adenovirus, ambas hebras contienen codificación de proteínas. Muchos de los mensajeros se sintetizan inicialmente en forma de un transcrito largo que proviene de dos tercios de la longitud del DNA. Este transcrito es modificado posteriormente.")
6
Nomenclatura de las hebras de DNA en la transcripción
Hebra molde Hebra menos (-) Hebra codante Hebra no molde Hebra más (+) Hebra no codante Las dos cadenas complementarias del DNA se definen por su función en la transcripción. El transcrito de RNA se sintetiza sobre la hebra molde complementaria (-) siendo idéntica en secuencia (con U en lugar de T) a la hebra codificante no molde (+).
 Hebra codante. Hebra no molde. Hebra más (+) Hebra no codante. Las dos cadenas complementarias del DNA se definen por su función en la transcripción. El transcrito de RNA se sintetiza sobre la hebra molde complementaria (-) siendo idéntica en secuencia (con U en lugar de T) a la hebra codificante no molde (+).")
7
Ejemplo de complementariedad
(5')CGCTATAGCTGTTT(3') Hebra de DNA no molde (+) (3')GCGATATCGACAAA(5') Hebra de DNA molde (-) (5')CGCUAUAGCUGUUU(3') Transcrito de RNA La hebra no molde, (+) también es llamada hebra sin-sentido. Esta presenta una secuencia idéntica al transcrito (sólo cambiando la timina por el uracilo en éste último). La hebra molde (-) es llamada hebra con sentido.
CGCTATAGCTGTTT(3 ) Hebra de DNA no molde (+) (3 )GCGATATCGACAAA(5 ) Hebra de DNA molde (-) (5 )CGCUAUAGCUGUUU(3 ) Transcrito de RNA. La hebra no molde, (+) también es llamada hebra sin-sentido. Esta presenta una secuencia idéntica al transcrito (sólo cambiando la timina por el uracilo en éste último). La hebra molde (-) es llamada hebra con sentido.")
8
RNA polimerasa Subunidad s Núcleo de la enzima
Estructura de subunidades de la RNA polimerasa de E. coli. Las sub- unidades (de la que hay dos) , ' y tienen masas moleculares relativas de 36,500, 151,000, 155,000, 11,000 y 70,000, respectivamente. La subunidad también se denomina 70. Se cree que el sitio catalítico para la síntesis de RNA se encuentra en la subunidad .
 , y tienen masas moleculares relativas de 36,500, 151,000, 155,000, 11,000 y 70,000, respectivamente. La subunidad también se denomina 70. Se cree que el sitio catalítico para la síntesis de RNA se encuentra en la subunidad .")
9
Promotores en E. coli. Resumen
Región -35 Espaciador Región -10 Inicio del RNA trp TTGACA N17 TTAACT N7 A tRNATyr TTTACA N16 TATGAT N7 A lac TTTACA N17 TATGTT N6 A Cinco promotores de E. coli. En ellos se encuentran promotores del operón de triptofano, lactosa y arabinosa. Las secuencias varían de un promotor a otro pero la comparación de muchos promotores pone de manifiesto similitudes en las regiones -10 y En la parte inferior se muestra la secuencias consenso de las regiones -10 y La región -10 se conoce como caja Pribnow (D. Pribnow, 1975). Todas las secuencias son de la hebra codante y se leen 5’-->3' de izquierda a derecha tal como es la convención en representaciones de este tipo. La región espaciadora contiene un número variable de nucleótidos. Sólo se muestra el primer nucleótidos que codifica el transcrito de RNA, posición +1. recA TTGATA N16 TATAAT N7 A araB, A, D CTGACG N18 TACTGT N6 A TTGACA TATAAT
. Todas las secuencias son de la hebra codante y se leen 5’-->3 de izquierda a derecha tal como es la convención en representaciones de este tipo. La región espaciadora contiene un número variable de nucleótidos. Sólo se muestra el primer nucleótidos que codifica el transcrito de RNA, posición +1. recA. TTGATA. N16. TATAAT. N7. A. araB, A, D. CTGACG. N18. TACTGT. N6. A. TTGACA. TATAAT.")
10
Promotores de E. coli. 1

11
Inhibidores de la transcripción
+ N H Acridina Sar L-Pro L-meVal D-Val L-Thr O C N NH2 CH3 Estructura molecular de la actinomicina D y de la acridina, inhibidores de la transcripción. La porción enmarcada en la actinomicina D es plana y se intercala entre dos pares de bases sucesivos GC en el DNA de doble cadena. Las dos estructuras peptídicas cíclicas de la molécula de la actinomicina D se unen al surco menor de la doble hélice. La sarcosina (Sar) es la N-metilglicina, meVal representa la metilvalina. las uniones entre sarcosina, L-prolina y D-valina son enlaces peptídicos. La acridina también actúa por intercalación en el DNA. Actinomicina D
 es la N-metilglicina, meVal representa la metilvalina. las uniones entre sarcosina, L-prolina y D-valina son enlaces peptídicos. La acridina también actúa por intercalación en el DNA. Actinomicina D.")
12
Terminación de la transcripción
Una vez que el proceso de transcripción ha comenzado la burbuja de transcripción avanza a medida que el RNA es transcrito y se continúa hasta que la polimerasa encuentra una secuencia que induce la disociación del complejo DNA-RNA y la separación de la enzima. En eucariontes esta secuencia no ha sido bien estudiada, mientras que para los procariontes como E. coli se han reconocido dos tipos de señales de terminación que se diferencian por la ausencia o presencia de un factor proteico que parece reconocer dicha señal.

13
Terminación rho independiente
Terminación independiente de rho (r), contiene una secuencia que induce a la formación de una horquilla en el transcrito de RNA de 10 a 15 nucleótidos antes del final, seguida por un tramo de poliA en la cadena molde que facilita la disociación de los elementos de la transcripción.
, contiene una secuencia que induce a la formación de una horquilla en el transcrito de RNA de 10 a 15 nucleótidos antes del final, seguida por un tramo de poliA en la cadena molde que facilita la disociación de los elementos de la transcripción.")
14
En la terminación dependiente de r no se presenta el tramo de poliA, aunque sí se forma una horquilla en la que la RNA polimerasa hace una pausa y si se encuentra la proteína r presente se detiene la transcripción. No se conoce a detalle el mecanismo de disociación, pero en este se produce la hidrólisis de ATP por efecto de r. Rho factor cannot bind to RNA coated with ribosomes, but Rho factor binds to specific regions in naked RNA. Rho factor then interacts with RNAP, causing it to fall off of the DNA which terminates transcription. Thus, when translation termination occurs within a gene it can cause transcriptional termination, preventing expression of downstream genes. This process is called translational polarity.

15
Regulación de la transcripción en procariontes.
Muchas proteínas bacterias son inducibles su síntesis es regulada dependiendo del estado nutricional de la célula. La expresión diferencial de los genes que codifican para esas proteínas esta regulada generalmente a nivel de la iniciación de la transcripción

16
Regulación 2 Basados en el modelo de Jacob y Monod del control transcripcional, la transcripción del operón lac, que codifica 3 proteínas inducibles, es reprimido por unión de la proteína represora de lac a la secuencia operadora. En presencia de lactosa o otros inductores, esta represión se levanta y el operón lac se transcribe.

17
Regulación 3 Las mutaciones en el promotor, que une a la RNA polimerasa, o en el operador, actúan en cis; esto es sólo afectan la expresión de los genes en la misma molécula de DNA en que ocurre la mutación.

18
Regulación 4 Las mutaciones en la secuencia de un operador que resulta en la disminución de la unión del represor, resultan en una transcripción constitutiva. Las mutaciones en una secuencia promotora, que afectan la afinidad de la unión de la RNA polimerasa pueden disminuir (mutación a la baja) o incrementar (mutación a la alta) la transcripción.
 o incrementar (mutación a la alta) la transcripción.")
19
Regulación 5 Los represores y los activadores actúan en trans; esto es, afectan a la expresión de los genes que regulan sin importar en qué molécula de DNA están localizados en la célula.

20
Las mutaciones Oc del Operón lac actúan en cis
Figure Experimental demonstration that Oc mutations are cis-acting. E. coli cells containing two copies of the lac operon are diagrammed. Diagonal lines indicate genes and control regions carrying mutations. In these cells, the lac operon on the bacterial chromosome has an Oc mutation in the operator and a mutation in the lacZ gene (lacZ-), which inactivates the β-galactosidase enzyme. The lac operon on the F-plasmid is wild-type. (Top) In the absence of the inducer IPTG, these cells constitutively express a functional lactose permease but no functional β-galactosidase. These results indicate that the Oc mutation only affects the genes on the same DNA (i.e., the chromosomal lac operon). If the Oc mutation were trans-acting, then transcription of the lacZ+ gene on the F-plasmid would have yielded observable β-galactosidase activity. (Bottom) In the presence of IPTG, both permease and β-galactosidase activity are observed. As diagrammed, transcription of the wild-type lacZ gene on the plasmid yields functional β-galactosidase.
, which inactivates the β-galactosidase enzyme. The lac operon on the F-plasmid is wild-type. (Top) In the absence of the inducer IPTG, these cells constitutively express a functional lactose permease but no functional β-galactosidase. These results indicate that the Oc mutation only affects the genes on the same DNA (i.e., the chromosomal lac operon). If the Oc mutation were trans-acting, then transcription of the lacZ+ gene on the F-plasmid would have yielded observable β-galactosidase activity. (Bottom) In the presence of IPTG, both permease and β-galactosidase activity are observed. As diagrammed, transcription of the wild-type lacZ gene on the plasmid yields functional β-galactosidase.")
21
Acción trans del Operón lac
Figure Experimental demonstration that the lacI+ gene is trans-acting. (Top) Cells carrying a single lacI- gene produce an inactive repressor; as a result, they express β-galactosidase and lactose permease constitutively. (Bottom) When a wild-type lacI+ gene is introduced into lacI- cells on a F-factor plasmid, the transformed cells produce functional repressor, which can bind to both lac operators. As a result, these cells do not express β-galactosidase or lactose permease in the absence of inducer.
 Cells carrying a single lacI- gene produce an inactive repressor; as a result, they express β-galactosidase and lactose permease constitutively. (Bottom) When a wild-type lacI+ gene is introduced into lacI- cells on a F-factor plasmid, the transformed cells produce functional repressor, which can bind to both lac operators. As a result, these cells do not express β-galactosidase or lactose permease in the absence of inducer.")
22
El inductor produce incremento en la transcripción del operón.
Figure Biochemical demonstration that inducer leads to an increase in lac operon transcription. Small samples of an E. coli culture growing in glucose medium were removed just before and at short intervals after addition of IPTG. [3H]uridine was added to each sample immediately after it was removed; after 20 seconds the cells were lysed and RNA was isolated. Each such pulse-labeled sample was hybridized to an excess of membrane-bound cloned lac DNA (see Figure 7-17). Radioactivity that is retained on the membrane is a measure of the rate of mRNA synthesis during the short pulse-labeling period. A plot of the percentage of total [3H]RNA that hybridized to lac DNA versus time after addition of IPTG indicates that the rate of mRNA synthesis increased rapidly after addition of IPTG, reaching a maximal rate in about 2 minutes. [Adapted from G. Contesse, M. Crepin, and F. Gros, 1970, in J. R. Beckwith and D. Zipser, eds., The Lactose Operon, Cold Spring Harbor Laboratory Press.]
. Radioactivity that is retained on the membrane is a measure of the rate of mRNA synthesis during the short pulse-labeling period. A plot of the percentage of total [3H]RNA that hybridized to lac DNA versus time after addition of IPTG indicates that the rate of mRNA synthesis increased rapidly after addition of IPTG, reaching a maximal rate in about 2 minutes. [Adapted from G. Contesse, M. Crepin, and F. Gros, 1970, in J. R. Beckwith and D. Zipser, eds., The Lactose Operon, Cold Spring Harbor Laboratory Press.]")
23
X-Gal 5-bromo-4-chloro-3-indolyl-b-D-galactoside

24
Regulación negativa en la transcripción
Operador (a) La molécula señal ( ) origina la disociación de la proteína reguladora del DNA DNA Promotor 5' 3' mRNA Los represores se unen a sitios específicos del DNA. Estos sitios se nombran operadores en procariotes. Los operadores están situados cerca o bien dentro del promotor de tal modo que la unión de la RNA polimeras o su movimiento a lo largo del DNA después de su unión, se bloquean siempre que el represor este presente. La regulación mediante una proteína represora se de-nomina regulación negativa. La unión del represor está regulada mediante una señal molecu-lar, generalmente una molécula pequeña y específica que se une al represor, induciendole un cambio conformacional. La inte-racción entre el represor y la molécula señal puede provocar tanto un aumento como una disminución de la transcripción. En algunos casos el cambio origna la disociación del represor unido al DNA del operador. En otros casos la interacción entre un represor inactivo y la molécula señal provoca la unión del represor al operador. (b) La molécula señal ( ) origina la unión de la proteína reguladora al DNA 5' 3' mRNA
 La molécula señal. ( ) origina la disociación. de la proteína reguladora. del DNA. DNA. Promotor. 5 3 mRNA. Los represores se unen a sitios específicos del DNA. Estos sitios se nombran operadores en procariotes. Los operadores están situados cerca o bien dentro del promotor de tal modo que la unión de la RNA polimeras o su movimiento a lo largo del DNA después de su unión, se bloquean siempre que el represor este presente. La regulación mediante una proteína represora se de-nomina regulación negativa. La unión del represor está regulada mediante una señal molecu-lar, generalmente una molécula pequeña y específica que se une al represor, induciendole un cambio conformacional. La inte-racción entre el represor y la molécula señal puede provocar tanto un aumento como una disminución de la transcripción. En algunos casos el cambio origna la disociación del represor unido al DNA del operador. En otros casos la interacción entre un represor inactivo y la molécula señal provoca la unión del represor al operador. (b) La molécula señal. ( ) origina la unión. de la proteína reguladora. al DNA. 5 3 mRNA.")
25
Regulación positiva en la transcripción
RNA polimerasa (a) La molécula señal ( ) origina la disociación de la proteína reguladora del DNA 5' 3' mRNA Los activadores son una alternativa molecular a los represores. La regulación mediada por activador se denomina regulación positiva. Los activadores se unen a sitios adyacentes al promotor reforzando tanto la unión como la actividad de la RNA polimerasa en ese promotor. Estos activadores se localizan gene-ralmente junto a promotores que tienen poca afinidad por la RNA polimerasa, por lo que la transcripción es insignificante sin el activador. Algunas veces el activador está unido normalmente al DNA y se disocia al unirse a la molécula señal. Cuando está unida al DNA, la proteína activadora facilita la unión de la RNA polimerasa y aumenta la tasa de inicio de la transcripción. En otros casos, el activador no se une al DNA hasta que no se encuentra unido a una señal molecular. La regulación positiva es habitual en eucariotas. (b) La molécula señal ( ) origina la unión de la proteína reguladora al DNA 5' 3' mRNA
 La molécula señal. ( ) origina la disociación. de la proteína reguladora. del DNA. 5 3 mRNA. Los activadores son una alternativa molecular a los represores. La regulación mediada por activador se denomina regulación positiva. Los activadores se unen a sitios adyacentes al promotor reforzando tanto la unión como la actividad de la RNA polimerasa en ese promotor. Estos activadores se localizan gene-ralmente junto a promotores que tienen poca afinidad por la RNA polimerasa, por lo que la transcripción es insignificante sin el activador. Algunas veces el activador está unido normalmente al DNA y se disocia al unirse a la molécula señal. Cuando está unida al DNA, la proteína activadora facilita la unión de la RNA polimerasa y aumenta la tasa de inicio de la transcripción. En otros casos, el activador no se une al DNA hasta que no se encuentra unido a una señal molecular. La regulación positiva es habitual en eucariotas. (b) La molécula señal. ( ) origina la unión. de la proteína reguladora. al DNA. 5 3 mRNA.")
26
Modelo general de operón
Sitio de unión del activador Sitio de unión del represor (operador) DNA Promotor A B C En un operón, los genes A, B y C se transcriben en un mRNA policistrónico. La secuencias reguladoras típicas incluyen sitios de unión para proteínas que pueden tanto activar como reprimir la transcripción a partir del promotor. Secuencias reguladoras
 DNA. Promotor. A. B. C. En un operón, los genes A, B y C se transcriben en un mRNA policistrónico. La secuencias reguladoras típicas incluyen sitios de unión para proteínas que pueden tanto activar como reprimir la transcripción a partir del promotor. Secuencias reguladoras.")
27
Operón lac reprimido PI I Z Y A P O Represor mRNA
El gen I codifica el represor Lac. Los genes lac S, Y y A codifican para la -galactosidasa, la galactósido permeasa y la transaceti- lasa, respectivamente. Los lugares P y O son el promotor y el operador, respectivamente, de los genes lac El lugar PI es el promotor del gen I. PI I Z Y A P O

28
Inducción del operón lac
PI I Z Y A P O DNA Alolactosa (o IPTG) El represor Lac sufre un cambio conformacional por la unión de la alolactosa. el represor se disocia del operador, permitiendo que proceda la transcripción. Otros -galactósidos como el isopropiltiogalactósido (IPTG), también pueden actuar como in-ductores. PI I Z Y A P O mRNA
 El represor Lac sufre un cambio conformacional por la unión de la alolactosa. el represor se disocia del operador, permitiendo que proceda la transcripción. Otros -galactósidos como el isopropiltiogalactósido (IPTG), también pueden actuar como in-ductores. PI. I. Z. Y. A. P. O. mRNA.")
29
Control positivo de la transcripción
Unlike the lac operon, the enhancer of the glnA gene is a little far from the promoter so that the bound activator does not immediately contact the polymerase. Nitrogen Regulatory Protein C (NTRC) can induce DNA looping to bring the activator in contact with the polymerase. Figure 4-D-5. The mechanism of transcriptional activation by NTRC. (a) The glnA gene is transcribed by the Sigma-54-containing polymerase which alone cannot initiate transcription. The unphosphorylated NTRC dimers can bind only one site at the enhancer, still insufficient to stimulate transcription. (b) The phosphorylated NTRC dimers can bind both sites of the enhancer. (c) Their binding induces DNA looping. Contact between the activator and the polymerase stabilizes the interaction between the polymerase and DNA, thereby initiating transcription. Una proteína reguladora promueve la unión de la RNA polimerasa, de manera que se incrementa la síntesis del mRNA.
 can induce DNA looping to bring the activator in contact with the polymerase. Figure 4-D-5. The mechanism of transcriptional activation by NTRC. (a) The glnA gene is transcribed by the Sigma-54-containing polymerase which alone cannot initiate transcription. The unphosphorylated NTRC dimers can bind only one site at the enhancer, still insufficient to stimulate transcription. (b) The phosphorylated NTRC dimers can bind both sites of the enhancer. (c) Their binding induces DNA looping. Contact between the activator and the polymerase stabilizes the interaction between the polymerase and DNA, thereby initiating transcription. Una proteína reguladora promueve la unión de la RNA polimerasa, de manera que se incrementa la síntesis del mRNA.")
30
Activación de la transcripción del operón lac
Sitio CAP 5'-ATTAATGTGAGTTAGCTCACTCATTAGGCACCCCA Región -35 Región -10 GGCTTTACACTTTATGCTTCCGGCTCGTATGTTGTGTG Sitio de unión a la RNA polimerasa El operón lac presenta otro mecanismo regulador cuando la cé- lula se halla en presencia simultanea de glucosa y lactosa. Ya que la primera es el metabolito de elección. El efeto represor de la glucosa está mediado por el cAMP y una proteína llamada proteína activadora del catabolito (CAP). La CAP es un homodímero con sitios de unión para el DNA y el cAMP. En ausencia de glucosa, la CAP se une a un lugar específico cerca- no al promotor lac incrementando la transcripción del RNA 50 veces. Por lo tanto CAP es un elemento de regulación positiva mientras que el represor de lac representa a la regulación ne-gativa. Los dos actuan simultaneamente; CAP tiene poco efecto en el sistema cuando el represor Lac bloquea la transcripción, y la disociación del represor del operador tiene poco efecto si CAP no está presente. Operador GAATTGTGAGCGTGATAACAATTTCACAC
. La CAP es un homodímero con sitios de unión para el DNA y el cAMP. En ausencia de glucosa, la CAP se une a un lugar específico cerca- no al promotor lac incrementando la transcripción del RNA 50 veces. Por lo tanto CAP es un elemento de regulación positiva mientras que el represor de lac representa a la regulación ne-gativa. Los dos actuan simultaneamente; CAP tiene poco efecto en el sistema cuando el represor Lac bloquea la transcripción, y la disociación del represor del operador tiene poco efecto si CAP no está presente. Operador. GAATTGTGAGCGTGATAACAATTTCACAC.")
31
Estructura tridimensional del homodímero CAP
Motivo de unión al DNA DNA Representación de CAP en cinta en la que la subunidades se muestran en blanco y azul. En rojo se muestra el motivo de unión al DNA hélice-giro-hélice. En azul obscuro las moléculas de cAMP. Representación de CAP (2 subunidades) unido al DNA, los pares de bases reconocidos por la proteína se muestran en verde. El DNA se curva alrededor de la proteína. cAMP
 unido al DNA, los pares de bases reconocidos por la proteína se muestran en verde. El DNA se curva alrededor de la proteína. cAMP.")
32
Efectos combinados de glucosa y lactosa en la expresión del operón lac
Deficiencia de lactosa Abundancia de lactosa P O Deficiencia de glucosa, abundancia de cAMP RNA polimerasa P O mRNA Sólo se dá una transcripción eficiente cuando la concentración de lactosa es alta y la de glucosa baja. P O Abundancia de glucosa, deficiencia de cAMP P O

33
Operón ara Activador Represor araC mRNA Sitio de unión de CAP araO2
L-Arabinosa Represor araC mRNA Sitio de unión de CAP araO2 araO1 araI El operón de arabinosa es un sistema de regulación más complejo que el de lac . Una única proteína efectúa un control positivo y negativo simultáneamente. Esta es la proteína AraC, y la unión a una molécula señal modifica su conformación, desde una forma represora que se une a una secuencia regula-dora de DNA hasta un activador que se une a una secuencia diferente. La proteína AraC regula su propia síntesis mediante la represión de la transcripción de su gen (autorregulación). Genes estructurales araC araB araA araD PC PBAD araBAD mRNA Sitios de Regulación
. Genes estructurales. araC. araB. araA. araD. PC. PBAD. araBAD mRNA. Sitios de Regulación.")
34
Regulación del operón ara (1)
Sitio de unión de CAP RNA polimerasa araO2 araI araC araBAD araO1 PC PBAD araC mRNA Cuando la proteína AraC es deficiente, su gen se transcribe desde su propio promotor. AraC regula su propia síntesis, uniendose a araO1 y reprimiento la transcripción del gen araC cuando su concentración supera las cuarenta copias por célula.

35
Regulación del operón ara (2)
Glucosa a niveles altos arabinosa a niveles bajos araO2 Proteínas AraC araC araO1 araBAD Cuando los niveles de arabinosa son bajos y los de glucosa altos, la proteína AraC unida tanto a araI como a araO2 forman un dímero, acercando estos dos sitios mediante la formación de un lazo de DNA de unos 210 pb. En esta conformación el sistema reprime la transcripción de los genes araBAD desde el promotor. Esta proteína se une también a araO1, reprimiendo la síntesis de AraC. PBAD araI PC Sitio de unión de CAP

36
Regulación del operón ara (3)
Glucosa a niveles bajos arabinosa a niveles altos CAP-cAMP RNA polimerasa Proteína AraC araC araBAD araO2 PBAD araI PC Cuando hay arabinosa y la concentración de glucosa es baja, la proteína AraC se une a la arabinosa y cambia su conformación para convertirse en un activador. Se abre el lazo de DNA, y la proteína AraC actúa conjuntamente con el complejo CAP-cAMP facilitando la transcripción. araBAD mRNA arabinosa

37
Operón de triptofano Región reguladora Genes estructurales Trp
Represor Trp Represor mRNA Guía (trpL) Atenuador trpR P O trpE trpD trpC trpB trpA El operón de triptofano de E. coli incluye cinco genes que codifican para las enzimas de la biosínteis del triptofano desde ácido corísmico. El mRNA sintetizado a partir de este operón tiene una vida media de sólo 3 min, permitiendo a la célula responder rápidamente frente a las necesidades variables de este aminoácido. El represor Trp es un homodímero en el que cada subunidad tiene 107 residuos de aminoácido. Cuando el triptofano es abundante, se une al represor Trp originando un cambio conformaional que permite al represor unirse a su ope-rón. El sito del operador trp se solapa con el del promotor, y la unión del represor bloquea la unión de la RNA pol. La velocidad de la síntesis de estas enzimas biosintéticas varia por un factor de 700 cuando el operón está activado. Cuando el sistema deja de estar reprimido, la velocidad de la transcripción se ajusta mediante un segundo proceso de regulación conocido como atenuación de la transcripción. Región reguladora Genes estructurales trp mRNA bajos niveles de triptofano mRNA atenuado altos niveles de triptofano
 Atenuador. trpR. P. O. trpE. trpD. trpC. trpB. trpA. El operón de triptofano de E. coli incluye cinco genes que codifican para las enzimas de la biosínteis del triptofano desde ácido corísmico. El mRNA sintetizado a partir de este operón tiene una vida media de sólo 3 min, permitiendo a la célula responder rápidamente frente a las necesidades variables de este aminoácido. El represor Trp es un homodímero en el que cada subunidad tiene 107 residuos de aminoácido. Cuando el triptofano es abundante, se une al represor Trp originando un cambio conformaional que permite al represor unirse a su ope-rón. El sito del operador trp se solapa con el del promotor, y la unión del represor bloquea la unión de la RNA pol. La velocidad de la síntesis de estas enzimas biosintéticas varia por un factor de 700 cuando el operón está activado. Cuando el sistema deja de estar reprimido, la velocidad de la transcripción se ajusta mediante un segundo proceso de regulación conocido como atenuación de la transcripción. Región reguladora. Genes estructurales. trp mRNA. bajos niveles. de triptofano. mRNA. atenuado. altos niveles de triptofano.")
38
Regulación del operón trp
Estructura atenuadora Péptido líder MKAIFVLKGWWRTS RNA polimerasa Ribosoma 4 3 1 2 5' UUU 3' mRNA Codones Trp DNA Péptido líder Cuando los niveles de triptofano son altos, el ribosoma traduce rápidamente la secuencia 1 (marco abierto de lectura codificante del péptido lider) y bloquea la secuencia 2 antes de que se transcriba la 3. La transcripción continua permitiendo que se pre-sente la atenuación con la estructura antiterminadora formada por las secuencias 3 y 4. Cuando los niveles de triptofano son bajos, el ribosoma se detie-ne en los codones Trp de la secuencia 1. La formación de estructuras apareadas entre las secuencias 2 y 3 evita la atenuación, debido a que la secuencia 3 no está disponible para formar la estructura atenuadora con la secuencia 4. MKAIFVLKG Genes estructurales 2 3 4 1 DNA RNA polimerasa
 y bloquea la secuencia 2 antes de que se transcriba la 3. La transcripción continua permitiendo que se pre-sente la atenuación con la estructura antiterminadora formada por las secuencias 3 y 4. Cuando los niveles de triptofano son bajos, el ribosoma se detie-ne en los codones Trp de la secuencia 1. La formación de estructuras apareadas entre las secuencias 2 y 3 evita la atenuación, debido a que la secuencia 3 no está disponible para formar la estructura atenuadora con la secuencia 4. MKAIFVLKG. Genes estructurales DNA. RNA. polimerasa.")
39
Secuencia guía de trpL Péptido guía M K A I F V L K G W W R T S - PARO
1 2 Secuencia del atenuador de triptofano. Se observa la secuencia del péptido guia y las posibles interacciones entre 1, 2, 3 y 4. 3 4 Polipéptido TrpE

40
Apareamiento del atenuador
La estructura del atenuador (apareamiento 3:4) provoca que la RNA polimerasa se detenga y termine la transcripcion. Por el contrario, al deternerse la traducción, el part 2:3 se forma impidiendo el apareamiento 3:4 y la RNA polimerasa no se detiene. Par 3:4 Par 2:3
 provoca que la RNA polimerasa se detenga y termine la transcripcion. Por el contrario, al deternerse la traducción, el part 2:3 se forma impidiendo el apareamiento 3:4 y la RNA polimerasa no se detiene. Par 3:4. Par 2:3.")
41
Otros mecanismos de atenuación
Las Bacteria Gram positivas (B. subtilis) utilizan la atenuación. El mecanismo es independiente de la traducción. Utiliza una proteína que se une al RNA cuando existe un efector. El efector puede ser el aminoácido regulado pero también su aminoacil-tRNA. La proteína se llama proteína trp de atenuación (en el caso del triptofano).
 utilizan la atenuación. El mecanismo es independiente de la traducción. Utiliza una proteína que se une al RNA cuando existe un efector. El efector puede ser el aminoácido regulado pero también su aminoacil-tRNA. La proteína se llama proteína trp de atenuación (en el caso del triptofano).")
42
Otros sistemas de atenuación
Operones de la biosíntesis de pirimidinas (E. coli traduccional y B. subtilis con proteína de atenuación). Atenuación traduccional. La traducción del péptido líder previene la traducción del siguiente gen en un mensajero policistrónico. Genes de resistencia a antibióticos en Gram positivas.
. Atenuación traduccional. La traducción del péptido líder previene la traducción del siguiente gen en un mensajero policistrónico. Genes de resistencia a antibióticos en Gram positivas.")
43
Otras redes de control global
Sistemas que regulan muchos genes simultáneamente en respuesta a una señal ambiental específica. Los sistemas de control global pueden incluir más de un regulón. Modulón es un grupo de genes que pueden responder a una proteína reguladora común incluso cuando son de diferentes regulones. Estimulón es un grupo de genes que responden a la misma señal ambiental

44
Factores sigma alternativos
Factor sigma es la subunidad de la RNA polimerasa que reconoce al promotor. La mayoría de genes de E. coli responden a s70. La cantidad del factor sigma alternativo regula la respuesta del E. coli sistema global. En E. coli hay 7 factores s diferentes. B. subtilis tiene un factor s más.

45
Algunos sistemas de control global por factor s en Escherichia coli.

46
Quorum sensing (percepción e quorum)
Señal regulatoria controlada por la densidad de población. Cada bacteria sintetiza una lactona de homoserina acilada (AHL). La AHL difunde hacia el exterior de la célula sin problema. Se alcanza un concentración intracelular alta de AHL cuando hay alta densidad celular. AHL se une a una proteína activadora y enciende genes. Sistema luciferasa bacteriana, genes de producción de biofilms (películas) en P. aeruginosa, en Staphylococcus aureus produce varias proteínas virulentas.
. La AHL difunde hacia el exterior de la célula sin problema. Se alcanza un concentración intracelular alta de AHL cuando hay alta densidad celular. AHL se une a una proteína activadora y enciende genes. Sistema luciferasa bacteriana, genes de producción de biofilms (películas) en P. aeruginosa, en Staphylococcus aureus produce varias proteínas virulentas.")
47
Transducción de señales y sistemas reguladores de dos componentes
La señal externa no es transmitida directamente a la proteína reguladora. La señal es detectada primero por un sensor La señal se transmite en una forma diferente hacia el sistema regulador en un proceso llamado transducción de señales.

48
Componentes del sistema regulador
Proteína detectora específica localizada en la membrana. Generalmente una quinasa detectora. Proteína reguladora de respuesta específica.

49
Quinasa detectora Enzima de membrana plamática que Fosforila compuestos Detecta la señal ambiental en la superficie externa y se fosforila a sí misma en un residuo de histidina Este fosfato se transmite a el regulador de respuesta intracelular. El regulador de respuesta intracelular es una proteína que controla la trascripción uniéndose al DNA.

50
Sistemas reguladores de dos componentes en la regulación de la transcripción de E. coli

51
Quimioaxis Movimiento de acercamiento o alejamiento bacteriano de un compuesto químico particular No es una respuesta a gradientes espaciales de una sustancia sino gradientes temporales Sienten el cambio en concentración de un compuesto fuera de la célula a intervalos Sistema de dos componentes para sentir este cambio y regulan el movimiento flagelar.

52
Proteínas involucradas en quimiotaxis
Proteínas sensoras: Aceptoras de grupos metilo (MCPs) En E. coli se han identificado cinco MCPs Todas son transmemebranales Las MCPs se unen directamente a un compuesto o indirectamente a una proteína periplasmática Se disparan una serie de reacciones que afectan la rotación del flagelo. Rotación antihorario, la célula continua, rotación horario, la bacteria da un viraje.
 En E. coli se han identificado cinco MCPs. Todas son transmemebranales. Las MCPs se unen directamente a un compuesto o indirectamente a una proteína periplasmática. Se disparan una serie de reacciones que afectan la rotación del flagelo. Rotación antihorario, la célula continua, rotación horario, la bacteria da un viraje.")
53
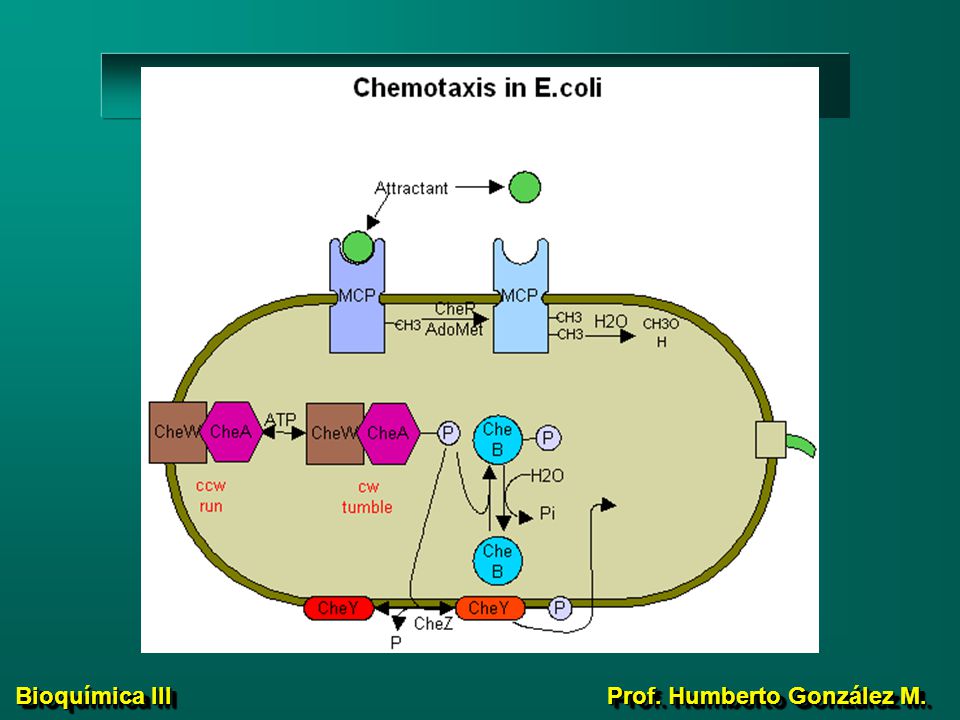
Quimiotáxis en E. coli CheW y CheA proteínas citoplásmicas,

55
La mayoría de sistemas de regulación controlan la transcripción mediante proteínas reguladoras.
Existen RNAs reguladores (antisentido)
")
56
Acoplamiento entre trascripción y traducción
RNA polimerasa DNA duplex 5' 3' 3' 5' Ribosoma mRNA 5' Dirección de transcripción mRNA En procariotes, el mRNA es traducido por los ribosomas cuando aún está siendo transcrito a partir del DNA por la RNA polimerasa. Esto es posible porque el mRNA procariótico no tiene que ser transportado de un núcleo al citoplasma antes de encontrar los ribosomas. En este esquema el ribosoma es dibujado mucho más pequeño que la RNA pol lo que no es cierto, pues el ribosoma es un orden de magnitud más grande que la enzima (ribosoma Mr 2.5 x 106 , RNApol 3.9 x 105). NH3+ NH3+ Dirección de traducción
. NH3+ NH3+ Dirección de traducción.")
57
Elementos reguladores lejanos en operones bacterianos
Figure Activation of sigma-54 -containing RNA polymerase at glnA promoter by NtrC. The polymerase binds to the glnA promoter, forming a closed complex, before being activated. In response to a low concentration of organic nitrogen, a protein kinase called NtrB phosphorylates dimeric NtrC (purple), which then binds to two enhancers (orange) centered at -108 and -140 from the transcription start site. The bound phosphorylated NtrC dimers interact with the bound σ54-polymerase, causing the intervening DNA to form a loop. The ATPase activity of NtrC then stimulates the polymerase to unwind the template strands at the start site, forming an open complex. Transcription of the glnA gene can then begin.
, which then binds to two enhancers (orange) centered at -108 and -140 from the transcription start site. The bound phosphorylated NtrC dimers interact with the bound σ54-polymerase, causing the intervening DNA to form a loop. The ATPase activity of NtrC then stimulates the polymerase to unwind the template strands at the start site, forming an open complex. Transcription of the glnA gene can then begin.")
59
Resumen de la transcripción

60
Splicing del gen de ovoalbúmina
F G L 1 2 3 4 5 6 7 DNA L 1 2 3 4 5 6 7 Transcrito primario A B C D E F G Cap Edición, rompimiento y poliadenilación Visión general de la modificación del mRNA de la ovoalbúmina (gen eucariótico). Los intrones están señalados con letras mientras que los exones están señalados con números. Alrededor de tres cuartas partes del RNA se eliminan durante la maduración. Los intrones pueden llegar a constituir más del 90% de la longitud en algunos genes. La RNA polimerasa II extiende el transcrito primario más allá del sitio de corte y poliadenilación. No se han definido las señales de terminación de la RNa polimerasa II. 7 intrones RNA extra 1 2 3 4 5 6 7 RNA maduro AAAAn 1.872 nucleótidos
. Los intrones están señalados con letras mientras que los exones están señalados con números. Alrededor de tres cuartas partes del RNA se eliminan durante la maduración. Los intrones pueden llegar a constituir más del 90% de la longitud en algunos genes. La RNA polimerasa II extiende el transcrito primario más allá del sitio de corte y poliadenilación. No se han definido las señales de terminación de la RNa polimerasa II. 7 intrones. RNA extra RNA maduro. AAAAn nucleótidos.")
61
Inicio Figure Transcription of DNA into RNA is catalyzed by RNA polymerase, which can initiate the synthesis of strands de novo on DNA templates. The nucleotide at the 5’ end of an RNA strand retains all three of its phosphate groups; all subsequent nucleotides release pyrophosphate (PPi) when added to the chain and retain only their α phosphate (red). The released PPi is subsequently hydrolyzed by pyrophosphatase to Pi, driving the equilibrium of the overall reaction toward chain elongation. In most cases, only one DNA strand is transcribed into RNA.
 when added to the chain and retain only their α phosphate (red). The released PPi is subsequently hydrolyzed by pyrophosphatase to Pi, driving the equilibrium of the overall reaction toward chain elongation. In most cases, only one DNA strand is transcribed into RNA.")
Presentaciones similares