Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Clase 1- Mecanismos epigenéticos

2
Epigenética Abarca las alteraciones heredables en la cromatina o la expresión génica, que no son acompañadas por cambios en la secuencia de ADN Otra definición: cambio en fenotipo que es heredable pero no involucra mutaciones en el ADN Principales mecanismos epigenéticos: Metilación de ADN Modificación post-traduccional de histonas Silenciamiento de genes mediado por ARN

3
Historia de Epigenética
Ligada al estudio de evolución y desarrollo Hasta 1950 la palabra se usaba de forma completamente diferente: todos los eventos del desarrollo que llevan de un cigoto fertilizado al organismo maduro En 1930 Muller describió una clase de mutaciones en Drosophila que consistían de translocaciones cromosómicas. Aún cuando todas las partes de la cromatina estaban representadas en la dosis correcta el fenotipo no era normal. Se debe a que estos rearreglos transladan a un gen a una región heterocromática

4
En algunas células el gen permanece activo, en otras se convierte en heterocromatina de ahí el patrón en el ojo

5
Historia Segundo aporte: estudios de desarrollo
Evidente que durante el desarrollo hay divergencia de fenotipos en las células y tejidos en diferenciación. Al parecer estos cambios podían ser heredados clonalmente por las células.

6
ADN es el mismo en todas las células somáticas del organismo
Los estudios de Avery, MacLeod y McCarty (1944) y Hershey y Chase (1952) demostraron que una molécula de ADN, sin proteínas, podía acarrear información genética Reforzado por la solución de Watson y Crick de la estructura del ADN (1953) Experimentos en Rana y Xenopus demostraron que la introducción de un núcleo de células embrionales tempranas en un oocito sin núcleo podía resultar en el desarrollo de un embrión Prueba de que las células del embrión todavía tenían todo el material genético Por lo tanto el programa de desarrollo y la especialización en expresión no es causada por cambios en el ADN.
 y Hershey y Chase (1952) demostraron que una molécula de ADN, sin proteínas, podía acarrear información genética. Reforzado por la solución de Watson y Crick de la estructura del ADN (1953) Experimentos en Rana y Xenopus demostraron que la introducción de un núcleo de células embrionales tempranas en un oocito sin núcleo podía resultar en el desarrollo de un embrión. Prueba de que las células del embrión todavía tenían todo el material genético. Por lo tanto el programa de desarrollo y la especialización en expresión no es causada por cambios en el ADN.")
7
Papel de la Metilación Inactivación del cromosoma X fue un modelo temprano para este tipo de inactivación Estaba claro que se inactivaba un cromosoma X al azar, y no había evidencia de cambios en la secuencia de ADN Riggs y Holliday (1975) y Pugh (1975) propusieron que la metilación de ADN podría actuar como una marca epigenética La base de su teoría es que diferentes enzimas se encargan de metilar ADN que no está metilado y ADN que ya tiene una banda metilada
 y Pugh (1975) propusieron que la metilación de ADN podría actuar como una marca epigenética. La base de su teoría es que diferentes enzimas se encargan de metilar ADN que no está metilado y ADN que ya tiene una banda metilada.")
8
Papel de la Metilación El primer evento es más difícil que el segundo
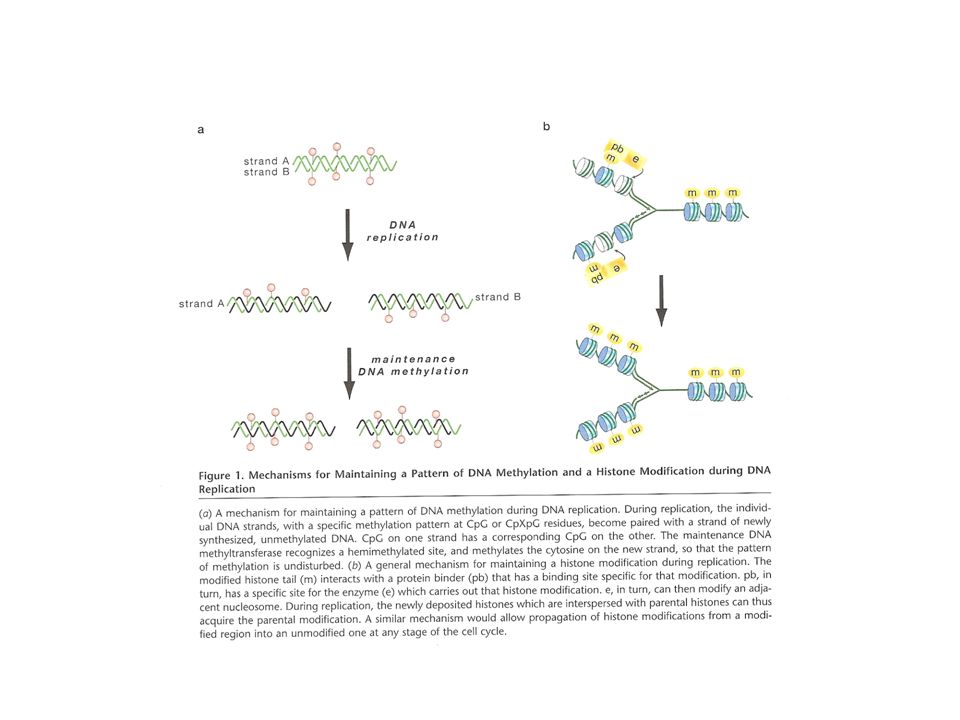
Cuando la primera banda se modifica, la banda complementaria es modificada rápidamente Una marca de metilación presente en la banda parental se copia a la banda hija después de la replicación, con el resultado de la transmisión del estado de metilación a la siguiente generación Poco después Bird (1978) aprovechó el hecho de que el principal blanco de metilación en animales es la secuencia CpG e introdujo el uso de enzimas de restricción sensibles a metilación para detectar el estado de metilación. Lo que se da es una propagación semiconservativa del patrón de metilación
 aprovechó el hecho de que el principal blanco de metilación en animales es la secuencia CpG e introdujo el uso de enzimas de restricción sensibles a metilación para detectar el estado de metilación. Lo que se da es una propagación semiconservativa del patrón de metilación.")
9
Papel de la Metilación En los años siguientes se ha enfocado mucha atención en: Los patrones endógenos de metilación La transmisión de esos patrones por la línea germinal El papel de metilación en silenciar la expresión de genes Mecanismos para la iniciación o inhibición de la metilación en un sitio previamente no metilado Identificación de enzimas responsables de metilación de novo y de mantener metilación en sitios metilados

10
Papel de la Metilación Sin embargo, estaba claro que este no podía ser el único mecanismo para la transmisión epigenética de la información Ej: el ejemplo en Drosophila no se puede explicar por metilación, porque Drosophila tiene niveles muy bajos de metilación de ADN

11
Papel de la cromatina Antes de que la mayoría de los estudios sobre metilación empezaran ya se había propuesto (Stedman y Stedman, 1950) que las histonas podrían actuar como represoras de la expresión génica Por un tiempo se creyó que las histonas eran proteínas supresoras que silenciaban la expresión génica en forma pasiva De acuerdo a esta visión, activar un gen consistía en elminar las histonas En 1964 Allfrey (1964) propuso que la acetilación de histonas podría estar correlacionada con la activación de genes y que la cromatina activa no necesariamente tendría que carecer de histonas
 que las histonas podrían actuar como represoras de la expresión génica. Por un tiempo se creyó que las histonas eran proteínas supresoras que silenciaban la expresión génica en forma pasiva. De acuerdo a esta visión, activar un gen consistía en elminar las histonas. En 1964 Allfrey (1964) propuso que la acetilación de histonas podría estar correlacionada con la activación de genes y que la cromatina activa no necesariamente tendría que carecer de histonas.")
12
Papel de la cromatina En 1974 Kornberg y Thomas descubrieron la estructura del nucleosoma, la subunidad básica de la cromatina A partir de 1980 y a lo largo de varios años Grunstein y colaboradores pudieron mostrar que las colas amino terminales de las histonas son esenciales para la regulación de la expresión génica y el establecimiento de dominios silenciosos en la cromatina Prueba última de conexión entre acetilación y expresión fue en 1996 cuando se descubrió que una acetiltransferasa de histonas de Tetrahymena era homóloga a la proteína regulatoria de la transcripción en levaduras: Gcn5 (Allis) Cómo es el funcionamiento: en los 90 se descubrió que tanto modificación de histonas como remodelación de nucleosomas están involucrados en preparar la cromatina para la transcripción
 Cómo es el funcionamiento: en los 90 se descubrió que tanto modificación de histonas como remodelación de nucleosomas están involucrados en preparar la cromatina para la transcripción.")
13
Papel de la cromatina Aún no estaba claro como la información del status de actividad se podía transmitir a través de la división celular Próximo paso: se descubrió que histonas modificadas reclutan proteínas que pueden afectar el estado funcional y estructural de la cromatina Ej: metilación de histona H3 resulta en reclutamiento de la proteína de heterocromatina HP1. HP1 recluta la enzima que hace esa metilación. Modelo: propagación progresiva del estado silenciado de la cromatina a lo largo de la región. Explica cómo se puede transmitir a través de la replicación

15
Mecanismos relacionados
Estos mecanismos explican cómo se mantiene el estado heterocromático No explican el establecimiento inicial de estructuras de silenciamiento de cromatina Evidencia a favor de papel de secuencias cortas de ARN, que se unen al ADN como parte de complejos que incluyen proteínas del grupo Polycomb y así inician la formación de heterocromatina Cambio epigenético que se transmite por meiosis y mitosis ha sido documentado en plantas, pero muy raramente en animales Curso: enfocado a transmisión mitótica de señales epigenéticas

16
Visión general y conceptos

17
Genética vs Epigenética
Uno de los descubrimientos más importantes en biología: estructura del ADN Permitió comprender la transmisión de información genética a nivel molecular

18
Genética vs Epigenética
Epigenética nace al tratar de explicar fenómenos „extraños“, no-Mendelianos Ej: inactivación X, expresión de un solo alelo, gemelos monocigóticos no completamente idénticos Tienen en común: secuencia de ADN no está alterada Reto de epigenética: explicar cómo se regula selectivamente uno de dos alelos en el núcleo, cómo se escoge y cómo se transmite esa escogencia

19
Modelos para estudio de epigenética
Drosophila Mamíferos Neurospora Maíz Arabidopsis Levadura

20
Fenómenos epigenéticos
Tienen en común que en eucariotas el ADN no está „desnudo“ ADN existe en un complejo como proteínas especializadas para formar la cromatina La cromatina se consideraba inicialmente como molécula pasiva cuya única función era el empaque y organización de ADN Sin embargo, existen diferentes formas de cromatina

21
Modificaciones a cromatina
Gran variedad de modificaciones post-traduccionales de histonas Remodelación de cromatina dependiente de energía, que mobiliza o altera estructuras de nucleosomas El movimiento dinámico de nuevas histonas (variantes) dentro y fuera de los nucleosomas Papel de ARNs no codificantes Metilación directa del ADN
 dentro y fuera de los nucleosomas. Papel de ARNs no codificantes. Metilación directa del ADN.")
22
Ahora se sabe que hay más

23
Modificaciones a cromatina
Muchas de las modificaciones son reversibles, poco probable que pasen por línea germinal Marcas transitorias son atractivas porque pueden crear cambios en cromatina en respuesta a estímulos internos y externos Algunas modificaciones de histonas (como metilación de lisina, metilación de ADN, y estructura alterada de nucleosomas), pueden permanecer por varias divisiones celulares Esto establece „estados epigenéticos“, o medios de lograr memoria celular: todavía no comprendidos
, pueden permanecer por varias divisiones celulares. Esto establece „estados epigenéticos , o medios de lograr memoria celular: todavía no comprendidos.")
24
Epigenomas El ADN es el mismo en todas las células del organismo
Este genoma va a ser epigenéticamente programado para generar „epigenomas“ diferentes en más de 200 tipos de célula Se ha propuesto que esto constituiría un „código epigenético“. Hipótesis: debe ser probada En términos modernos (moleculares) epigenética sería: „la suma de las alteraciones al templete de cromatina que colectivamente establecen y propagan patrones diferentes de expresión génica y silenciamiento del mismo genoma“
 epigenética sería: „la suma de las alteraciones al templete de cromatina que colectivamente establecen y propagan patrones diferentes de expresión génica y silenciamiento del mismo genoma")
25
La cromatina Nucleosoma: unidad repetitiva fundamental de la cromatina
Compuesto por: un octámero de proteínas que contiene dos moléculas de cada una de 4 tipos de histona (H2A, H2B, H3 y H4), alrededor de las cuales hay 147 pb de ADN arrollados Histonas en el corazón del nucleosoma son pequeñas y altamente básicas Tienen un dominio globular y „colas de histona“ flexibles y poco estructuradas, que sobresalen de la superficie del nucleosoma
, alrededor de las cuales hay 147 pb de ADN arrollados. Histonas en el corazón del nucleosoma son pequeñas y altamente básicas. Tienen un dominio globular y „colas de histona flexibles y poco estructuradas, que sobresalen de la superficie del nucleosoma.")
26
Nucleosoma

27
Histonas Secuencia de aa altamente conservada de levadura a humanos
Alta conservación sugiere que su función es crítica, incluso en las colas Las colas, particularmente de H3 y H4 van a estar sujetas a extensas modificaciones post-traduccionales Acetilación y metilación de histonas, especialmente, H3 y H4 fueron de las primeras modificaciones covalentes en ser descritas y se propuso que correlacionaban con cambios positivos y negativos en actividad transcripcional

28
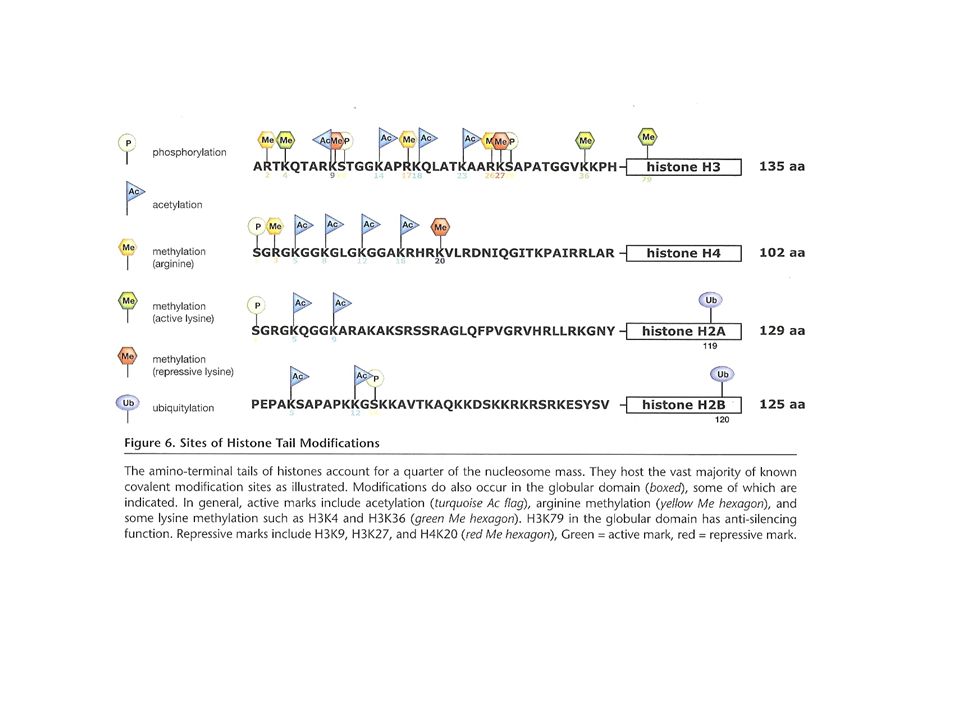
Histonas modificadas en las colas

29
Modificaciones covalentes de histonas
Acetilación Metilación Fosforilación Ubiquitinación Sumoilación (unión de un péptido, SUMO, similar a ubiquitina) ADP-ribosilación Biotinilación Isomerización de prolina
 ADP-ribosilación. Biotinilación. Isomerización de prolina.")
31
Modificaciones covalentes de histonas
Enzimas específicas y complejos enzimáticos catalizan la formación de estas marcas En ciertas regiones de la cromatina, los nucleosomas pueden contener proteínas variantes de histona, en lugar de histonas „core“ Esto contribuye a marcar las regiones de los cromosomas para funciones especializadas Se conocen variantes de H2A y H3, pero aún no para H2B y H4

33
Organización de la cromatina
Cromatina: polímero de ADN-nucleosomas es una molécula dinámica que existe en muchas configuraciones Tradicionalmente se clasificaba como eucromatina o heterocromatina basado en tinción Eucromatina: cromatina descondensada. Puede estar activa o inactiva transcripcionalmente. Heterocromatina: cromatina altamente condensada y silenciada Si está silenciada permanentemente: heterocromatina constitutiva, los genes se expresarán muy rara vez Si está silenciada sólo en algunos momentos del desarrollo o CC: heterocromatina facultativa

34
Organización de la cromatina
La fibra de cromatina no siempre consiste de nucleosomas a intervalos regulares como se aprende (11nm): ADN periódicamente rodeando unidades repetitivas de nucleosomas
: ADN periódicamente rodeando unidades repetitivas de nucleosomas.")
35
Organización de la cromatina
Nucleosomas pueden estar empacados en forma irregular y formar estructuras de un mayor orden Estas conformaciones diferentes de la cromatina ocurren en diversas regiones del genoma durante la especificación del destino de una célula o en diferentes momentos del CC

36
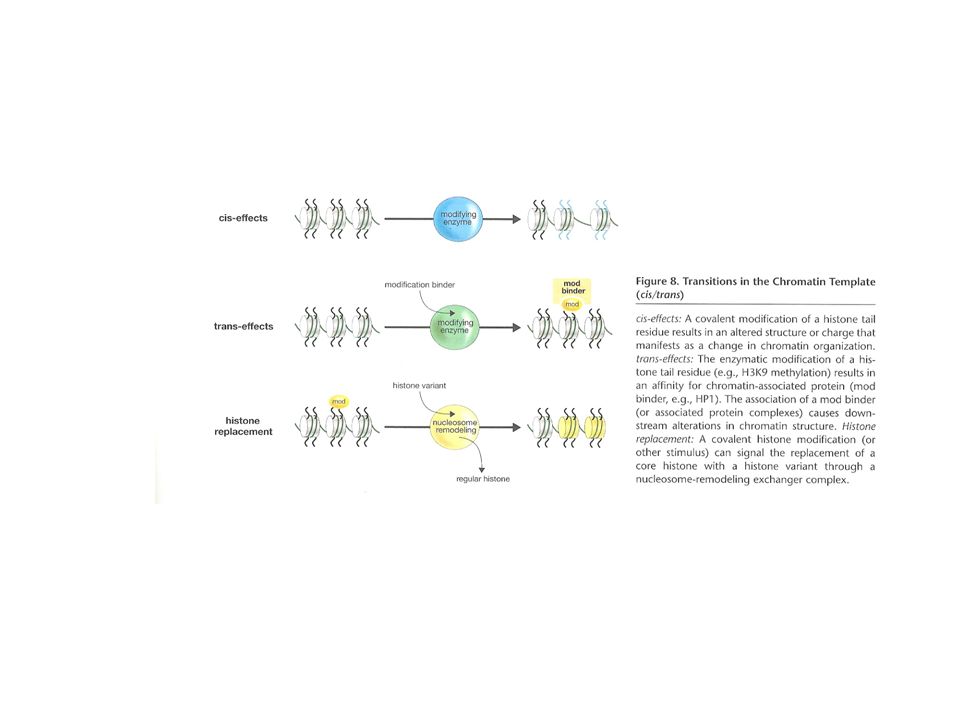
Efectos cis y trans El arreglo de los nucleosomas en el templete de 11 nm puede ser alterado por efectos cis y trans de las colas de histonas modificadas covalentemente Efectos cis: causados por cambios en las propiedades físicas de las colas modificadas de las histonas, como modulación en la carga electroestática o la estructura de la cola, que afecta los contactos entre nucleosomas Ej:acetilación de histonas. Se cree que neutraliza cargas positivas de las colas altamente básicas de las histonas, generando expansión localizada de la fibra de cromatina y, por lo tanto, permitiendo mejor acceso de maquinaria de transcripción al ADN

37
Efectos cis y trans Efectos trans de modificación de colas de histonas: reclutar elementos de modificación-unión a la cromatina Algunos elementos de unión tienen afinidad particular por ciertas colas de histona. Funcionan como el „velcro“ que une el polímero de cromatina con un complejo enzimático Ej: el bromodominio-que reconoce residuos de histonas acetilados- es a menudo parte de una acetil transferasa de histonas (HAT) que acetila histonas, como parte de un complejo grande de remodelación de cromatina
 que acetila histonas, como parte de un complejo grande de remodelación de cromatina.")
38
Efectos cis y trans Otro ej trans: modificaciones de histonas de la cola o el núcleo pueden atraer complejos de remodelación dependientes de ATP a la fibra de 11nm que se requieren para la transición de eucromatina en „pausa“ a un estado transcripcionalmente activo Estos remodeladores de cromatina usan la E para causar cambios en los contactos ADN-histona, que resulta en movimiento de los nucleosomas Estas son interacciones no-covalentes

40
Organización cromatina
Estructuras de cromatina más compactas y represivas se logran al reclutar la histona „linker“ H1 y/o factores „arquitectónicos“ asociados a cromatina como proteína de heterocromatina 1 (HP1) o Polycomb (PC) HP1 tiene 8 isoformas: posiblemente con diferentes funciones Existe debate sobre la organización de la forma de 30 nm: estudios actuales sugieren un arreglo en zig zag de ADN „linker“ que conecta dos torres de partículas de nucleosomas Organización a un mayor nivel de la cromatina no ha sido elucidada
 o Polycomb (PC) HP1 tiene 8 isoformas: posiblemente con diferentes funciones. Existe debate sobre la organización de la forma de 30 nm: estudios actuales sugieren un arreglo en zig zag de ADN „linker que conecta dos torres de partículas de nucleosomas. Organización a un mayor nivel de la cromatina no ha sido elucidada.")
41
Condensación en metafase
La forma más condensada del ADN se observa en metafase. Permite segregación de los cromosomas Implica reestructuración del ADN de una molécula de 2m a cromosomas con un diámetro promedio de 1,5 um. Se compacta veces. Se logra por hiperfosforilación del linker H1 y la histona „core“ H3 y la acción ATP-dependiente de los complejos de condensina y cohesina y topoisomerasa II.

42
Telómeros y centrómeros
Importantes en la dinámica de cromosomas Telómeros: puntas del cromosoma, dan protección y solucionan la replicación de los finales de moléculas de ADN Centrómeros: lugar de anclaje de los microtúbulos del huso mitótico en la división nuclear Ambas regiones son fundamentales en la segregación de cromosomas Ambas se pueden distinguir de eucromatina y también de otras regiones heterocromáticas por la presencia de estructuras únicas de la cromatina que son altamente represivas para la actividad de genes y la recombinación

43
Telómeros y centrómeros
El mover algún gen de su lugar original en eucromatina a un lugar cerca de un centrómero o telómero causa que el gen no se exprese Centrómeros y telómeros tienen marcas características como histonas hipoacetiladas El ensamblaje correcto de la heterocromatina del centrómero es crítica para la mitosis y meiosis La marca del centrómero es epigenética: no importa tanto la secuencia de ADN sino las modificaciones

44
Eucromatina y Heterocromatina
Eucromatina es la cromatina „activa“ y consiste principalmente de secuencias codificantes (<4% del genoma en mamíferos) Evidencia de que eucromatina tiene una configuración „abierta“, lo que hace que esté lista para la expresión de genes pero no necesariamente transcripcionalmente activa Algunos genes son housekeeping, otros regulados por programa de desarrollo o el ambiente Expresión va a depender de elementos cis (promotor, enhancers) y trans (FT) Pregunta es si cromatina tiene el papel de hacer las secuencias blanco accesibles en cada tipo de célula
 Evidencia de que eucromatina tiene una configuración „abierta , lo que hace que esté lista para la expresión de genes pero no necesariamente transcripcionalmente activa. Algunos genes son housekeeping, otros regulados por programa de desarrollo o el ambiente. Expresión va a depender de elementos cis (promotor, enhancers) y trans (FT) Pregunta es si cromatina tiene el papel de hacer las secuencias blanco accesibles en cada tipo de célula.")
45
Eucromatina y Heterocromatina
En las cercanías de los promotores, regiones ricas en AT, a menudo no hay nucleosomas, lo que promueve la ocupación por factores de transcripción Pero: unión de FT no es suficiente para asegurar transcripción Existe una maquinaria que remodela nucleosomas, con modificaciones activadoras (como acetilación y metilación de H3K4) y facilita la transcripción Después de transcripción para llegar a ARN maduro se requiere splicing, etc. Término eucromatina involucra una maquinaria compleja
 y facilita la transcripción. Después de transcripción para llegar a ARN maduro se requiere splicing, etc. Término eucromatina involucra una maquinaria compleja.")
46
Eucromatina y Heterocromatina
96% del genoma son secuencias no-codificantes y repetitivas Para bloquear la transcripción en la heterocromatina: Deacetilación de colas de histonas Metilación de residuos específicos de lisina Reclutamiento de proteínas asociadas a histonas (ej. HP1) Establecimiento de metilación de ADN Evidencia sugiere que heterocromatina es el estado „default“
 Establecimiento de metilación de ADN. Evidencia sugiere que heterocromatina es el estado „default")
47
Eucromatina y Heterocromatina
¿Qué funciones puede tener la heterocromatina? Definición del centrómero: crítico para división celular Heterocromatina constitutiva en los telómeros protege los finales de los cromosomas Defensa contra ADN invasor

48
Modificacione a histonas y el código de histonas
Modificación actúan como tipo de „switch“: encendido/apagado Acetilación de histonas asociada con dominios activos, permisivos para transcripción Ciertos residuos de histonas fosforilados: cromatina condensada Modificaciones hechas por complejos enzimáticos Muchas de estas enzimas catalizan reacciones con alta especificidad (dependiendo de factores internos o externos)
")
49
Complejos enzimáticos
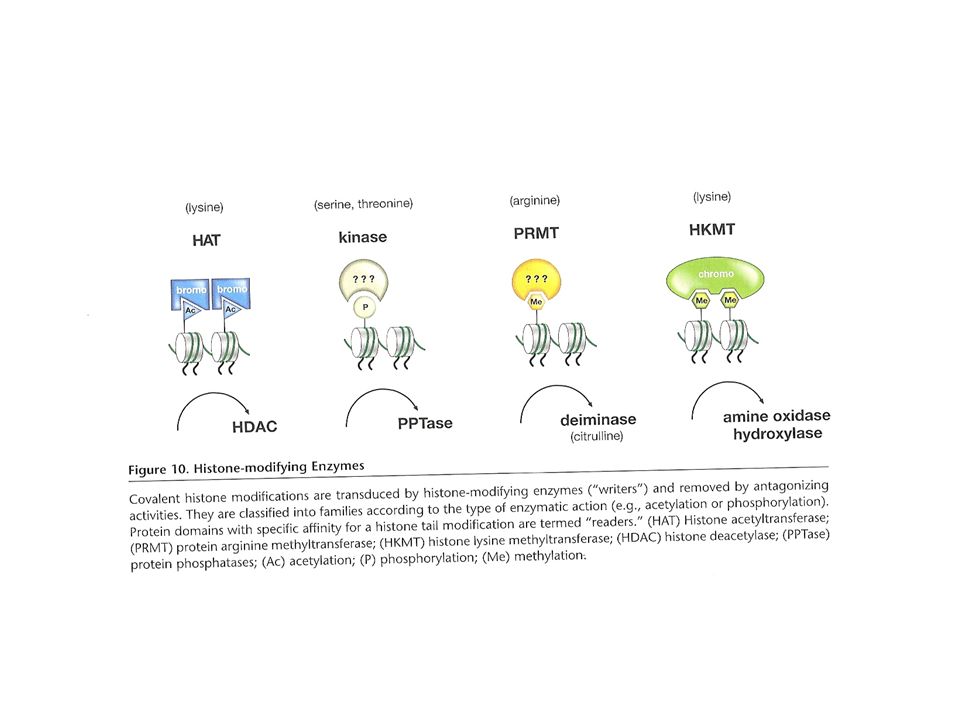
Acetilasas de histonas (HAT): Acetilan residuos específicos de lisina. Su efecto es revertido por la acción de deacetilasas de histonas (HDACs) Quinasas de histonas: fosforilan residuos específicos de Ser o Thr y las fosfatasas eliminan marcas de fosforilación Quinasas mitóticas (como CDK): catalizan la fosforilación de histonas „core“ (H3) o „linker“ (H1)
: Acetilan residuos específicos de lisina. Su efecto es revertido por la acción de deacetilasas de histonas (HDACs) Quinasas de histonas: fosforilan residuos específicos de Ser o Thr y las fosfatasas eliminan marcas de fosforilación. Quinasas mitóticas (como CDK): catalizan la fosforilación de histonas „core (H3) o „linker (H1)")
50
Complejos enzimáticos
Enzimas metiladoras, 2 clases: Metiltransferasas de arginina (PRMTs) Metiltransferasas de lisina de histonas (HKMTs) Metilación de arginina revertida por deiminasas Residuos de lisina metilados son químicamente más estables y parecen poder propagarse en las divisiones celulares Metilación de lisina revertida por demetilasa específica de lisina (LSD1)
 Metiltransferasas de lisina de histonas (HKMTs) Metilación de arginina revertida por deiminasas. Residuos de lisina metilados son químicamente más estables y parecen poder propagarse en las divisiones celulares. Metilación de lisina revertida por demetilasa específica de lisina (LSD1)")
52
Modificaciones de histonas no ocurren en forma aislada

53
Código de histonas No está claro si algo así existe
Aún si existiera, no es universal Sin embargo, estudios de todo el genoma si detectan patrones Ej: fuerte correlación de H3K4me3 con promotores activos y de metilación de H3K9 con heterocromatina en „silencio“ Una modificación no conduce inevitablemente a un resultado

54
Complejos de remodelación de cromatina
Estos complejos usan E (hidrólisis de ATP) para cambiar la composición de cromatina y nucleosomas en forma no-covalente Los nucleosomas, sobre todo si están rodeados por factores represivos de cromatina, imponen a menudo una inhibición a la maquinaria de transcripción Sólo algunos FT y reguladores (pero no la maquinaria basal de transcripción) pueden ganar acceso a sus sitios de unión Este problema es solucionado por complejos de proteínas que mueven los nucleosomas y/o alteran su estructura Complejos remodeladores de cromatina a menudo actúan en conjunto con enzimas activadores modificadoras de cromatina
 para cambiar la composición de cromatina y nucleosomas en forma no-covalente. Los nucleosomas, sobre todo si están rodeados por factores represivos de cromatina, imponen a menudo una inhibición a la maquinaria de transcripción. Sólo algunos FT y reguladores (pero no la maquinaria basal de transcripción) pueden ganar acceso a sus sitios de unión. Este problema es solucionado por complejos de proteínas que mueven los nucleosomas y/o alteran su estructura. Complejos remodeladores de cromatina a menudo actúan en conjunto con enzimas activadores modificadoras de cromatina.")
55
Complejos de remodelación de cromatina
Dos familias principales SNF2H o ISWI Brahma o SWI/SNF Familia SNF2H/ISWI mueve los nucleosomas a lo largo del ADN Familia Brahma/SWI/SNF altera la estructura del nucleosoma de forma transitoria, y así expone contactos ADN-histona

57
Variantes de Histonas Otras actividades que usan E actúan como complejos de reemplazo, que reemplazan histonas convencionales „core“ con variantes especializadas de histonas O reemplazan histonas modificadas por histonas no modificadas Podría ser una manera de hacer „borrón y cuenta y nueva“

58
Variantes de histonas Además de la iniciación de la transcripción, la presencia de los nucleosomas también obstruye la elongación por parte de la ARN pol II Una serie de modificaciones de histonas y efectores actúan en conjunto con los complejos de remodelación de cromatina para permitir el paso de la pol II por los nucleosomas Variantes de histonas le dan una composición diferente a la cromatina que prepara a las regiones cromosómicas para diferentes funciones Ej: en genes transcripcionalmente activos H3 es reemplazada por H3.3. Reemplazo de H2A con H2A.Z correlacionado con actividad transcripcional e indica el extremo 5‘ de promotores libres de nucleosomas

59
Variantes de histonas Contrario a la creencia generalizada de que las histonas se sintetizan en la fase S del ciclo celular, la síntesis y sustitución de muchas de estas variantes de histonas ocurre independientemente de la replicación Por lo tanto se puede dar acción rápida: p.ej en respuesta a daños en el ADN

60
Metilación de ADN Primer mecanismo en ser correlacionado con represión génica Consiste en agregar un grupo metilo a residuos de citosina en el ADN Ocurre en dinucleótidos CpG en mamíferos ADN metilado enriquecido en regiones no codificantes y elementos repetitivos, pero no en las islas CpG de genes activos Metilación sirve como un mecanismo de defensa para silenciar genomas de origen extraño (ej. Secuencias virales

61
Metilación de ADN Metiltransferasas de ADN (DNMTs) son los efectores de la metilación Pueden catalizar la metilación de novo o el mantenimiento de la metilación en ADN hemimetilado después de la replicación Pérdida de la habilidad de mantener metilación resulta en enfermedades como ICF, pérdida en regulación de niveles de metilación : cáncer ¿Qué señales dirigen a DNMTs a metilar ciertas regiones de ADN? Secuencias altamente repetitivas (centrómero) usan la marca represiva de metilación de H3K9 para dirigir la metilación de novo Immunodeficiency, centromeric instability and facial abnormalities
 usan la marca represiva de metilación de H3K9 para dirigir la metilación de novo. Immunodeficiency, centromeric instability and facial abnormalities.")
62
Metilación de ADN No completamente claro como la metilación reprime la expresión Probablemente efecto en trans Methyl-CpG-binding domain proteins (MBD) se unen a sitios metilados Ej: una MBD, la MeCP2 se une a CpGs metilados y recluta deacetilasas de histonas (HDAC) para mediar la formación de marcas represivas en las histonas
 se unen a sitios metilados. Ej: una MBD, la MeCP2 se une a CpGs metilados y recluta deacetilasas de histonas (HDAC) para mediar la formación de marcas represivas en las histonas.")
63
Metilación de ADN Tanto en plantas como mamíferos placentados (pero no marsupiales) los loci imprinted tienen ADN metilado: es el mecanismo común en la evolución para estabilizar represión génica Regiones altamente repetitivas que están metiladas se vuelven mutagénicas cuando pierden metilación y causan inestabilidad genómica global: cáncer Pero: citosinas individuales metiladas tienden a mutar espontáneamente a T. Se cree que esto es beneficioso como defensa contra secuencias parasíticas de ADN como transposones
 los loci imprinted tienen ADN metilado: es el mecanismo común en la evolución para estabilizar represión génica. Regiones altamente repetitivas que están metiladas se vuelven mutagénicas cuando pierden metilación y causan inestabilidad genómica global: cáncer. Pero: citosinas individuales metiladas tienden a mutar espontáneamente a T. Se cree que esto es beneficioso como defensa contra secuencias parasíticas de ADN como transposones.")
64
RNAi y represión de genes mediada por ARN
Papel de heterocromatina en centrómero y telómero en integridad genómica: reconsiderar la clasificación como „ADN basura“ ¿Es posible que esas secuencias no sean completamente “silenciosas”? Experimentos en levadura unieron dos áreas de investigación: RNAi y las modificaciones características de heterocromatina En el exp. Mutaciones de cuaquier componente de la maquinaria de RNAi causaba defectos en la segregación de cromosomas. Esto se debe a la incapacidad de estabilizar la heterocromatina centromérica e hizo evidente el papel de mecanismos mediados por RNAi para producir dominios silenciosos de heterocromatina y la importancia de la heterocromatina en mantener la integridad del genoma

65
RNAi

66
RNAi y represión de genes mediada por ARN
En mamíferos también: tratamiento de células mamarias permeabilizadas con RNasa A elimina las marcas heterocromáticas H3K9me3 Si se eliminan factores de procesamiento de siRNA en vertebrados se afecta la metilación de H3K9 y la unión de HP1 a heterocromatina pericentromérica Interesante y no explicado: células madre embrionarias sin maquinaria RNAi proliferan pero no se diferencian

67
Polycomb y Tritorax Dentro de los principales efectores que pueden transducir señales a la cromatina y participan en mantener la identidad celular (dan memoria celular) están los grupos de genes polycomb (PcG) y tritorax (trxG) Reguladores clave de proliferación celular e identidad celular en eucariotas multicelulares Involucrados en cascadas de señales que responden a mitógenos y morfógenos
 están los grupos de genes polycomb (PcG) y tritorax (trxG) Reguladores clave de proliferación celular e identidad celular en eucariotas multicelulares. Involucrados en cascadas de señales que responden a mitógenos y morfógenos.")
68
Polycomb y Tritorax Los dos grupos de proteínas funcionan en forma esencialmente antagónica La familia PcG establece un estado silenciado de la cromatina La familia trxG por lo general propaga actividad génica Primera evidencia molecular de memoria celular o epigenética: identificación del gen Pc que estabiliza patrones de represión por varias generaciones celulares PC tiene un dominio similar a HP1 que se une a histonas

69
Polycomb y Tritorax Se han identificado aprox. 20 genes PcG y 17 trxG en Drosophila Genes PcG codifican: Proteínas de unión al ADN Enzimas modificadoras de histonas Otros factores represivos asociados a cromatina Genes trxG codifican: Factores de transcripción Enzimas remodeladoras de cromatina dependientes de ATP (ej. Brahma) HKMT (ej: familia MLL) Mecanismo por el cual complejos con PcG o trxG se dirigen a regiones de cromatina reguladas en el desarrollo no es aún conocido
 HKMT (ej: familia MLL) Mecanismo por el cual complejos con PcG o trxG se dirigen a regiones de cromatina reguladas en el desarrollo no es aún conocido.")
70
Polycomb y Tritorax En Drosophila la represión génica heredable requiere que se reclute complejos de PcG a elementos de ADN llamados elementos de respuesta polycomb (PRE) No se ha logrado identificar secuencias equivalentes en humanos El mecanismo no está claro: a menudo los PRE están a kilobases del inicio de transcripción del gen blanco
 No se ha logrado identificar secuencias equivalentes en humanos. El mecanismo no está claro: a menudo los PRE están a kilobases del inicio de transcripción del gen blanco.")
71
Polycomb y Tritorax Mantienen estado activo de expresión génica y revierten (o previenen) el silenciamiento mediado por PcG Menos claro aún que funcionamiento de PcG Cambios de cromatina documentados: incorporación de marcas „activas“ de metilación de lisina por HKMTs y la lectura de esas marcas Aumento en niveles de trxG: riesgo aumentado de cáncer de próstata, de mama, mieloma múltiple y leucemia Aumento de marcas represivas de histonas (p.ej. por exceso de PcG) se asocia con senescencia
 se asocia con senescencia.")
72
Heterocromatina facultativa
Silenciamiento mediado por PcG y la inactivación del cromosoma X son ejemplos de transiciones reguladas en el desarrollo entre los estados activos e inactivos de la cromatina Esto contrasta con heterocromatina constitutiva (como en los dominios pericentroméricos) La heterocromatina facultativa ocurre en regiones codificantes del genoma, donde el silenciamiento depende de decisiones del desarollo que especifican un destino celular específico (puede a veces ser reversible)
 La heterocromatina facultativa ocurre en regiones codificantes del genoma, donde el silenciamiento depende de decisiones del desarollo que especifican un destino celular específico (puede a veces ser reversible)")
73
Reprogramación del destino celular
La célula germinal y las células embrionales tempranas se distinguen de otras por su totipotencia innata Aunque hay unos 200 tipos de células en mamíferos, hay dos transiciones principales de diferenciación De célula pluripotente (célula madre) a célula completamente diferenciada Entre estado de reposo (G0) y proliferación
 a célula completamente diferenciada. Entre estado de reposo (G0) y proliferación.")
74
Reprogramación del destino celular
La regeneración del hígado y reparación de células musculares son excepciones Por lo general tejidos en mamíferos no se pueden reprogramar Para poder clonar es necesaria la reprogramación del epigenoma para poder introducirlo en el oocito y que empiece de 0

75
Reprogramación del destino celular
Obstáculos a reprogramación en mamíferos: Algunas marcas epigenéticas se transmiten establemente en las divisiones celulares y se resisten a la reprogramación Un núcleo de célula somática no puede recapitular la asimetría de marcas epigenéticas en un embrión fertilizado (parte de la madre, parte del padre) Al transferir al núcleo al oocito se modifican los loci imprinted que son claves en desarrollo fetal y de la placenta La mayoría de embriones clonados son abortados Impronta epigenética perturbada: podría ser la causa de la baja eficiencia del clonaje y de técnicas de reproducción asistida
 Al transferir al núcleo al oocito se modifican los loci imprinted que son claves en desarrollo fetal y de la placenta. La mayoría de embriones clonados son abortados. Impronta epigenética perturbada: podría ser la causa de la baja eficiencia del clonaje y de técnicas de reproducción asistida.")
76
Control epigenético Muy rara vez va a haber correlación entre una modificación a cromatina y un estado epigenético Más bien el estado epigenético es generado por la combinación y efecto acumulado de varios factores (factores de unión al ADN, modificación histonas, variantes de histonas, remodelación de nucleosomas, metilación de ADN y ARN no codificante) Unión de FT es por lo general transitoria. Se requieren en cada división celular subsecuente. El control epigenético puede potenciar una señal primaria a generaciones celulares sucesivas (pero no indefinidas) por la transmisión heredable de información por el templete de cromatina
 Unión de FT es por lo general transitoria. Se requieren en cada división celular subsecuente. El control epigenético puede potenciar una señal primaria a generaciones celulares sucesivas (pero no indefinidas) por la transmisión heredable de información por el templete de cromatina.")
77
Control epigenético Permite generar memoria celular a mediano plazo
Enzimas modificadoras a menudo requieren cofactores (ATP, acetil coA, SAM), cuyos niveles están dictados por ambiente (ej. dieta) Debido a que modificaciones de histonas probablemente se originaron como respuesta a estímulos externos
, cuyos niveles están dictados por ambiente (ej. dieta) Debido a que modificaciones de histonas probablemente se originaron como respuesta a estímulos externos.")
Presentaciones similares









 fue un proyecto de investigación científica con el objetivo fundamental de determinar la secuencia de.>")











